Строение ладонь: российские учёные создали технологию идентификации по строению сети венозных сосудов на руке — РТ на русском
российские учёные создали технологию идентификации по строению сети венозных сосудов на руке — РТ на русском
Российские разработчики создали технологию идентификации человека по индивидуальному строению сети венозных сосудов ладони. По их утверждению, венозный «рисунок» окончательно формируется к 12 годам и не меняется в течение жизни человека, поэтому он, как и папиллярный узор на пальцах, может быть способом биометрической идентификации. Подобную систему можно использовать в офисах, аэропортах и на вокзалах, на производствах, в спортивных центрах, банках, в розничных сетях, на предприятиях общественного питания и даже для доступа к автотранспорту и частным домам, заявляют специалисты.
Российские разработчики создали технологию идентификации человека по индивидуальному строению сети венозных сосудов ладони. Об этом RT сообщила пресс-служба «Платформы НТИ» (Национальной технологической инициативы). Разработчик — екатеринбургская компания Biosmart.
Проект сканера ладони Palmjet использует венозный «рисунок».
«Подобную систему можно использовать в офисах, аэропортах и на вокзалах, на производствах, в спортивных центрах, банках, в розничных сетях, на предприятиях общественного питания и даже для доступа к автотранспорту и частным домам», — отметил в беседе с RT пресс-секретарь «Платформы НТИ» Юрий Сушинов.
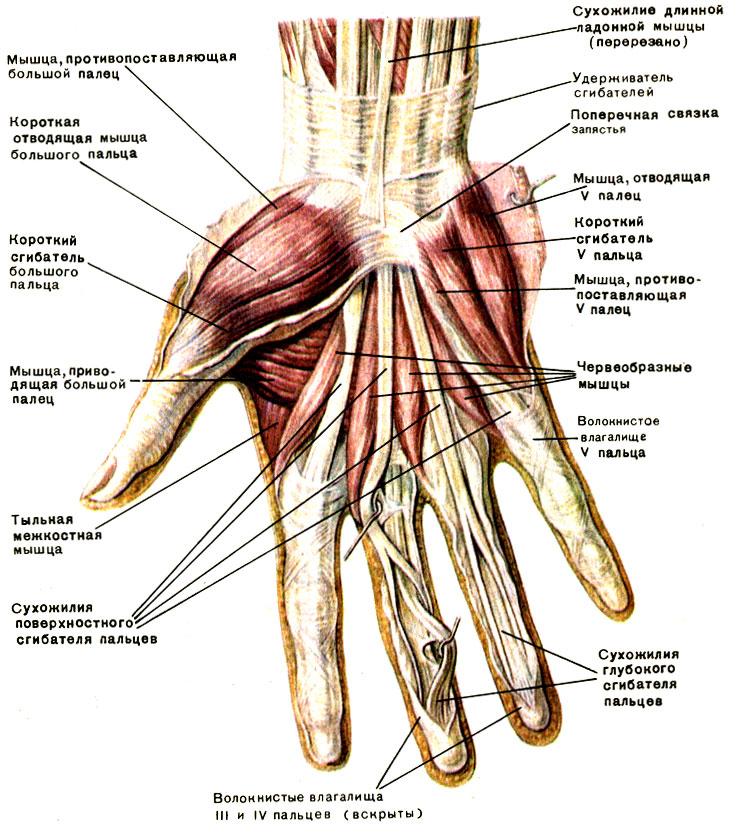
По словам разработчиков, подобные сканеры обладают мультифункциональными возможностями. Они позволяют избегать подмены электронных пропусков, а также могут вести учёт прогулов и опозданий сотрудников. Устройства сканируют ладонь в мультиспектральном инфракрасном свете и считывают её отражение.
«Гемоглобин в венах поглощает часть ИК-излучения, поэтому на отражении проявляется узор кровеносных сосудов. Математические алгоритмы преобразуют узор в цифровой код и упаковывают его в зашифрованный файл-шаблон размером всего 4 килобайта», — объясняют авторы проекта.
Также по теме
«Подготовиться к возможным потрясениям»: IT-специалист — о перспективах и рисках в сфере искусственного интеллектаИскусственному интеллекту нельзя давать право принимать решения, связанные с человеческими судьбами, а для контроля над ним необходимы…
Как рассказали RT специалисты, такой биометрический шаблон сохраняется в базе данных. Чтобы идентифицировать человека, устройство сканирует его ладонь и сравнивает с файлами из базы. За это отвечает искусственный интеллект — нейросетевой алгоритм. Весь процесс идентификации, уверяют разработчики, занимает менее полсекунды.
По их данным, состояние поверхности руки не имеет значения: сканер вен распознает человека как по грязной или влажной ладони, так и ладони с порезами, мозолями и даже в тонкой медицинской перчатке. Прикасаться к поверхности при проверке не обязательно — сканер фиксирует рисунок вен с расстояния 3—9 см.
«Из-за пандемии бесконтактные методы идентификации будут очень активно расти, и мы наблюдаем это на примере лицевой биометрии, но и у бесконтактного сканера вен ладони есть хорошие шансы стать лидером рынка», — сообщила в беседе с RT PR-директор компании-разработчика Анна Маликова.
В планах специалистов занять от 25 до 30% российского рынка биометрии.
Фиброаденома
Фиброаденома – это доброкачественная опухоль молочной железы. Пальпаторно фиброаденома (ФА) определяется как болезненное (жгучая, колющая боль), легко смещаемое объемное образование плотной консистенции, округлой или овальной формы. Средние размеры ФА обычно варьируют от 0,3 до 1,5 см и, как правило, не превышают 3 см в диаметре. Опухоль имеет гладкую поверхность, ровные, волнистые или крупнобугристые контуры, которые четко ограничены и не спаяны с окружающими тканями. Кожные покровы пораженной молочной железы не изменены.
Согласно данным различных источников литературы, ФА диагностируется у 10% женского населения и дебютирует в различные возрастные интервалы, но чаще – в периоды репродуктивной активности (в среднем с 16 до 40 лет).
Основным этиологическим фактором ФА является гормональный дисбаланс, а именно гиперэстрогенемия с одновременным снижением уровня прогестерона. Т.е. наибольший риск развития или усиленного роста ФА возникает во время гормональных перестроек организма (половое созревание, беременность, лактация, предменопаузальный и менопаузальный периоды, сопровождающиеся заместительной гормональной терапией, опухоли щитовидной железы, патологические процессы в печени и пр.). ФА состоит из двух видов тканей: фиброзная (соединительная) и эпителиальная.
В зависимости от соотношения этих тканей ФА классифицируют на 4 типа: периканаликулярные, интраканаликулярные, смешанные и филлоидные (листовидные) фиброаденомы. Риск малигнизации (озлокачествления, перехода в саркому молочной железы) у ФА I — III типов отсутствует в отличие от IV типа — листовидной ФА. Листовидная ФА молочной железы – это фиброэпителиальная опухоль, развивающаяся из внутрипротоковой фиброаденомы. Патогномоничным признаком данного типа ФА является быстрый рост опухолевого образования. Это как бы промежуточная форма между саркомой и фиброаденомой. По причине перечисленных особенностей, а, главное, повышенного риска малигнизации, листовидные ФА требуют немедленного оперативного вмешательства.
Риск малигнизации (озлокачествления, перехода в саркому молочной железы) у ФА I — III типов отсутствует в отличие от IV типа — листовидной ФА. Листовидная ФА молочной железы – это фиброэпителиальная опухоль, развивающаяся из внутрипротоковой фиброаденомы. Патогномоничным признаком данного типа ФА является быстрый рост опухолевого образования. Это как бы промежуточная форма между саркомой и фиброаденомой. По причине перечисленных особенностей, а, главное, повышенного риска малигнизации, листовидные ФА требуют немедленного оперативного вмешательства.
Также необходимо отметить, что ФА может иметь множественный характер и рецидивирующее течение. Кроме того, известны клинические случаи обратного развития ФА. Обычно это небольших размеров опухоли, подвергшиеся длительной консервативной терапии.
Факторы, диктующие необходимость хирургического лечения ФА:
- Быстрый рост опухоли, а именно увеличение в размерах объемного образования в два раза в течение 3–4 месяцев.
- Наличие множественных (в т.
 ч. двусторонних) ФА.
ч. двусторонних) ФА. - Большие размеры опухоли, деформирующие окружающие ткани молочной железы.
- Наличие выраженной или атипической пролиферации эпителия в опухоли (по результатам цитологического исследования при пункционно-аспирационной биопсии).
- Беременность – как гормональный дисбаланс, инициирующий рост опухолевой ткани. Увеличенная в размерах ФА механически сдавливает млечные протоки, что затрудняет лактацию и способствует развитию патологических процессов в молочной железе (например, лактостаза или мастита).
- Филлоидная (листовидная опухоль).
 ч. двусторонних) ФА.
ч. двусторонних) ФА.Алгоритм диагностических действий при наличии опухолевого образования в молочной железе (в том числе, непальпируемой формы).
- Консультация онколога – маммолога.
- УЗИ молочных желез и региональных лимфатических узлов (в т.ч. с эластографией — дополнительная технология, используемая во время традиционного УЗИ, которая дает возможность оценить эластичность тканей исследуемого органа). Дело в том, что все ткани человека, в том числе и поврежденные тем или иным патологическим процессом, имеют определенную эластичность (жесткость).
- Маммография (прицельная и дополнительные рентгенограммы интересующей зоны в прямой, боковой, либо косой проекциях).
- Пункционная биопсия или трепан-биопсия (диагностическая пункция под рентген или УЗ – контролем, выполняемая с целью верификации диагноза и определения распространенности опухолевого процесса).
- Предоперационная маркировка опухоли при непальпируемой форме (накожная маркировка или чрезкожная установка маркировочной иглы «гарпун» под УЗ- или рентген-контролем).
 Дело в том, что все ткани человека, в том числе и поврежденные тем или иным патологическим процессом, имеют определенную эластичность (жесткость).
Дело в том, что все ткани человека, в том числе и поврежденные тем или иным патологическим процессом, имеют определенную эластичность (жесткость).Объем хирургического вмешательства, выполняемого по поводу ФА молочной железы:
- Энуклеация – вылущивание (enucleo – вынимать ядро, очищать от скорлупы). Применяется при отсутствии подозрений на наличие злокачественного процесса. При этом дефект тканей молочной железы (как осложнение операции) минимален или отсутствует вообще.
- Секторальная резекция молочной железы – удаление опухолевого образования вместе с окружающими тканями молочной железы с последующим косметическим дефектом различной степени выраженности, который устраняется различными методами маммопластики (с использованием эндопротеза или местными тканями).
- Подкожная мастэктомия с одномоментной реконструкцией груди эндопротезом (имплантом) или местными тканями (при больших размерах опухоли или множественных ФА).
В нашем центре синтезированы общехирургические и косметические принципы при выборе методов в хирургии молочной железы. Разработаны и внедрены операции, которые обеспечивают не только адекватную радикальность, но и превосходный эстетический результат.
Контрактура Дюпюитрена — Сеть МЦ «Доктор Боголюбов»
Контрактура Дюпюитрена (ладонный фиброматоз) – невоспалительное заболевание, в процессе которого происходит рубцовое перерождение ладонных сухожилий.
Контрактура Дюпюитрена – избыточное развитие соединительной ткани в области сухожилий сгибателей одного или нескольких пальцев. Процесс локализуется на ладони. Развивается постепенно, возникает по невыясненным причинам. Приводит к ограничению разгибания и формированию сгибательной контрактуры одного или нескольких пальцев.
Контрактура Дюпюитрена – достаточно распространенное в ортопедии и травматологии заболевание, которое чаще наблюдается у мужчин среднего возраста. В половине случаев носит двухсторонний характер. Примерно в 40% случаев поражается безымянный палец, в 35% — мизинец, в 16% — средний палец, в 2-3% — первый и второй палец. У женщин выявляется в 6-10 раз реже и протекает более благоприятно. При возникновении в молодом возрасте характерно более быстрое прогрессирование.
Контрактура Дюпюитрена не связана с нарушениями белкового, углеводного или солевого обмена. Некоторые авторы утверждают, что наблюдается определенная связь между возникновением болезни и сахарным диабетом, однако эта теория пока не доказана.
Существуют также травматическая (вследствие травмы), конституционная (наследственные особенности строения ладонного апоневроза) и неврогенная (поражение периферических нервов) теории, однако мнения ученых остаются противоречивыми. В пользу конституционной теории свидетельствует наследственная предрасположенность. В 25-30% случаев у больных есть близкие кровные родственники, страдающие тем же заболеванием.
В пользу конституционной теории свидетельствует наследственная предрасположенность. В 25-30% случаев у больных есть близкие кровные родственники, страдающие тем же заболеванием.
Симптомы
Контрактура Дюпюитрена имеет очень характерную клиническую картину, которую трудно спутать с симптомами других заболеваний. На ладони больного выявляется уплотнение, образованное узлом и одним или несколькими подкожными тяжами. Разгибание пальца ограничено.
Первым признаком развития контрактуры Дюпюитрена обычно становится уплотнение на ладонной поверхности кисти, обычно – в области пястно-фаланговых суставов IV-V пальцев. В последующем плотный узелок медленно увеличивается в размере. Появляются тяжи, отходящие от него к основной, а затем – и к средней фаланге пораженного пальца. Из-за укорочения сухожилия сначала формируется контрактура в пястно-фаланговом, а потом — и в проксимальном (расположенном ближе к центру тела) межфаланговом суставе.
Кожа вокруг узла становится более плотной и постепенно спаивается с подлежащими тканями. Из-за этого в области поражения появляются выпуклости и втяжения. При попытке разогнуть палец узел и тяжи становятся более четкими, хорошо видимыми.
Обычно контрактура Дюпюитрена формируется без болей и только около 10% пациентов предъявляют жалобы на более или менее выраженный болевой синдром. Боли, как правило, отдают в предплечье или даже плечо.
Для контрактуры Дюпюитрена характерно прогрессирующее течение. Скорость прогрессирования болезни может колебаться и не зависит от каких-то внешних обстоятельств.
Степени
С учетом выраженности симптомов выделяют три степени контрактуры Дюпюитрена:
Первая. На ладони обнаруживается плотный узелок диаметром 0,5-1 см. Есть тяж, расположенный на ладони или достигающий области пястно-фалангового сустава. Иногда выявляется болезненность при пальпации.
Вторая. Тяж становится более грубым и жестким, распространяется на основную фалангу. Кожа также грубеет и спаивается с ладонным апоневрозом. В области поражения возникают видимые воронкообразные углубления и втянутые складки. Пораженный палец (или пальцы) согнуты в пястно-фаланговом суставе под углом 100 градусов, разгибание невозможно.
Третья. Тяж распространяется на среднюю, реже – на ногтевую фалангу. В пястно-фаланговом суставе выявляется сгибательная контрактура с углом 90 или менее градусов. Разгибание в межфаланговом суставе ограничено, степень ограничения может варьироваться. В тяжелых случаях фаланги расположены под острым углом друг к другу. Возможен подвывих или даже анкилоз.
Скорость прогрессирования контрактуры Дюпюитрена трудно прогнозировать. Иногда незначительное ограничение сохраняется на протяжении нескольких лет или даже десятилетий, а иногда от появления первых симптомов до развития тугоподвижности проходит всего несколько месяцев. Возможен также вариант с длительным стабильным течением, которое сменяется быстрым прогрессированием.
Возможен также вариант с длительным стабильным течением, которое сменяется быстрым прогрессированием.
Диагностика
Диагноз контрактура Дюпюитрена выставляется на основании жалоб пациента и характерной клинической картины. В ходе осмотра врач пальпирует ладонь пациента, выявляя узлы и тяжи, а также оценивает амплитуду движений в суставе.
Дополнительные лабораторные и инструментальные исследования для подтверждения диагноза обычно не требуются.
Лечение
Лечением контрактуры Дюпюитрена занимаются травматологи и ортопеды. Лечение может быть как консервативным, так и оперативным. Выбор методов производится с учетом выраженности патологических изменений.
Консервативная терапия применяется на начальных стадиях конрактуры Дюпюитрена. Пациенту назначают физиолечение (тепловые процедуры) и специальные упражнения для растяжения ладонного апоневроза. Могут также использоваться съемные лонгеты, фиксирующие пальцы в положении разгибания. Как правило, их надевают в ночное время, а днем снимают.
Могут также использоваться съемные лонгеты, фиксирующие пальцы в положении разгибания. Как правило, их надевают в ночное время, а днем снимают.
При упорном болевом синдроме используются лечебные блокады с гормональными препаратами (дипроспан, кеналог, гидрокортизон и т.д.). Раствор препарата смешивают с местным анестезирующим средством и вводят в область болезненного узла. Обычно эффект одной блокады сохраняется в течение 6-8 недель. Следует учитывать, что использование гормонов относится к числу методов лечения, которые следует применять с осторожностью.
Консервативные средства не могут устранить все проявления болезни. Они лишь замедляют скорость развития контрактуры. Единственным радикальным способом лечения остается хирургическая операция.
Четкие рекомендации по поводу выраженности симптомов, при которых необходимо проводить оперативное лечение, в настоящее время отсутствуют. Принятие решения о хирургическом вмешательстве основывается на скорости прогрессирования заболевания и жалобах пациента на боли, ограничение движений и связанные с этим трудности при самообслуживании или выполнении профессиональных обязанностей.
Обычно врачи рекомендуют оперативное вмешательство при наличии сгибательной контрактуры с углом 30 или более градусов. Целью операции, как правило, является иссечение рубцово измененной ткани и восстановление полного объема движений в суставах. Однако в тяжелых случаях, особенно – при застарелых контрактурах пациенту может быть предложен артродез (создание неподвижного сустава с фиксацией пальца в функционально выгодном положении)
Реконструктивная операция по поводу контрактуры Дюпюитрена может проводиться под общим наркозом или местной анестезией. При выраженных изменениях со стороны кожи и ладонного апоневроза хирургическое вмешательство бывает достаточно длительным, поэтому в таких случаях рекомендуется общий наркоз.
Существует множество вариантов разреза при контрактурах Дюпюитрена. Наиболее распространен поперечный разрез в области ладонной складки в сочетании с L- или S-образными разрезами по ладонной поверхности основных фаланг пальцев. Выбор конкретного способа производится с учетом особенностей расположения рубцовой ткани.
Выбор конкретного способа производится с учетом особенностей расположения рубцовой ткани.
В ходе операции ладонный апоневроз полностью или частично иссекают.
Затем рана ушивается и дренируется резиновым выпускником. На ладонь накладывается тугая давящая повязка, которая препятствует скоплению крови и развитию новых рубцовых изменений. Руку фиксируют лонгетой так, чтобы пальцы находились в функционально выгодном положении. Швы обычно снимают на десятый день. В последующем пациенту назначают лечебную гимнастику для восстановления объема движений в пальцах.
Иногда (особенно – при раннем возникновении и быстром прогрессировании) в течение нескольких лет или десятилетий может возникнуть рецидив контрактуры. В этом случае требуется повторная операция.
По всем вопросам лечения и диагностики Контрактуры Дюпюитрена вы можете обратится к нашим врачам травматологам ортопедам
Почему не получаются красивые стрелки?
В видеоуроках и пошаговых фотографиях подробно показывают, как рисовать идеальные стрелки и даже дают полный чек-лист средств. А они всё равно смотрятся не так гармонично, почему? Как правило, такой макияж показывают на моделях с классическим открытым подвижным веком, с хорошей кожей и другими “удобными” предпосылками. Возможно, этот вид стрелки и способ её нанесения вам не подходят, потому и ожидания расходятся с результатом.
А они всё равно смотрятся не так гармонично, почему? Как правило, такой макияж показывают на моделях с классическим открытым подвижным веком, с хорошей кожей и другими “удобными” предпосылками. Возможно, этот вид стрелки и способ её нанесения вам не подходят, потому и ожидания расходятся с результатом.
Что не так
1. Во внешнем уголке века складки и морщинки. Дело необязательно в возрасте – таким может быть просто генетическое строение глаза. В таком случае любая, даже самая старательно нарисованная графичная стрелка будет ломаться и ещё больше подчёркивать морщинки. Такому типу глаза нужна мягкая стрелка с растушевкой. Кстати, этот способ самый простой, не требует много времени и ювелирной точности. Наберите на кисточку карандаш и наметьте ею хвостик при открытом глазе. Затем пройдитесь по ресничному краю с середины века и выровняйте стрелку. Кисть для этого нужна упругая, с коротким ворсом (кисточка для теней не подходит), а карандаш – мягкий каял. Закрепите теневым скульптором, можно прозрачной пудрой, но скульптор создаст мягкую дымку. Накрасьте ресницы, тогда стрелку за ними будет не видно, но макияж станет выразительнее.
Затем пройдитесь по ресничному краю с середины века и выровняйте стрелку. Кисть для этого нужна упругая, с коротким ворсом (кисточка для теней не подходит), а карандаш – мягкий каял. Закрепите теневым скульптором, можно прозрачной пудрой, но скульптор создаст мягкую дымку. Накрасьте ресницы, тогда стрелку за ними будет не видно, но макияж станет выразительнее.
2. Нависшее веко. Под тяжёлым веком стрелку практически не видно. Таким глазам тоже подходит растушёванная стрелка. Ещё лучше добавить растушёванные тени сверху и снизу.
3. Неподходящее средство. Для чёткой графичной стрелки нужен лайнер или фломастер. Для мягкой стрелки подходит гелевый карандаш. Он устойчивый, хорошо держится на жирной коже, но плохо растушёвывается, потому что быстро застывает. Для растушёванной стрелки – мягкий карандаш или кремовая подводка. Подводить слизистую, прорисовывать межресничное пространство, делать макияж в стиле смоки нужно тоже с карандашом каял. Для графичной стрелки нужна тонкая и плоская кисть со скошенным краем. Для мягкой растушёванной нужна упругая с коротким ворсом. Кисть для теней не подходит.
Для графичной стрелки нужна тонкая и плоская кисть со скошенным краем. Для мягкой растушёванной нужна упругая с коротким ворсом. Кисть для теней не подходит.
Чёткая графичная стрелка (фломастер или подводка):
Мягкая стрелка (гелевый карандаш):
Растушёванная стрелка (карандаш кайал и кисть):
4. Близко посаженные глаза. С таким типом стрелку нужно начинать вести от середины и ни коем случае не обводить чёрным внутренний угол.
5. Широко посаженные глаза. Не удлинять стрелку к виску, а вот обвести внутренний уголок глаз, наоборот, желательно.
6. Карандаш “течёт” и отпечатывается на веке. Всегда запечатывайте макияж рассыпчатой прозрачной пудрой, сухим скульптором или тенями в цвет карандаша. Пользоваться только пушистой кистью и набирать небольшое количество средства. Стряхнуть через ладонь, а на кожу нанести лишь лёгкую вуаль.
Стряхнуть через ладонь, а на кожу нанести лишь лёгкую вуаль.
Если вам всё ещё не поддалось искусство рисования идеальных стрелок, попробуйте новые методы и разные варианты косметики: кисти, карандаши или подводки.
Миличкина Алёна
Как выбрать детские перчатки и варежки
Содержание статьи
1. Стандартные размеры
2. Как подобрать зимний аксессуар в интернет-магазине
3. Выбор детских перчаток по возрасту
4. Полезные советы
Детская кожа, особенно на руках, нуждается в особой защите и уходе. Ситуация обостряется зимой, когда ребенок постоянно норовит поиграть в снежки, покататься с горки или начать лепить снежную бабу. Защитить руки от ветра и низких температур помогают варежки и перчатки. Для наибольшего комфорта они должны быть изготовлены из теплых влагостойких материалов, иметь удобную конструкцию и главное – соответствовать анатомическим особенностям кисти. В нашей статье вы узнаете, как выбрать размер детских перчаток и варежек, особенно если вы оформляете покупку в онлайн-магазине.
Защитить руки от ветра и низких температур помогают варежки и перчатки. Для наибольшего комфорта они должны быть изготовлены из теплых влагостойких материалов, иметь удобную конструкцию и главное – соответствовать анатомическим особенностям кисти. В нашей статье вы узнаете, как выбрать размер детских перчаток и варежек, особенно если вы оформляете покупку в онлайн-магазине.
Стандартные размеры
До того как начать выбирать размер детских варежек, нужно понять, по какому принципу он определяется. Российские производители при этом ориентируются на мерку, снятую с самой широкой части ладони, но не захватывающую большой палец. Причем, согласно российским стандартам, размер равен этой мерке. То есть если обхват ладони составляет 13 см, то и размер будет 13.
При покупке толстых зимних перчаток или варежек можно выбирать аксессуары на 1-2 размера больше. Так ребенок сможет свободно двигать пальцами и согревать их за счет небольшой воздушной «подушки». Однако изделие не должно быть слишком просторным, иначе в нем будет скапливаться холодный воздух.
Однако изделие не должно быть слишком просторным, иначе в нем будет скапливаться холодный воздух.
Как подобрать зимний аксессуар в интернет-магазине
Как правило, интернет-магазин, в котором представлена детская одежда и аксессуары, прикрепляет внизу каталога или на странице конкретной модели размерную таблицу и инструкцию по снятию мерок. На иностранных онлайн-площадках указываются международные размеры, на российских – российские и международные. Некоторые сайты в качестве ориентира берут возраст ребенка.
Чтобы измерить параметры кисти, понадобится только сантиметровая лента. Далее нужно:
1. Обхватить самую широкую часть ладони доминирующей руки (у правшей – правая, у левшей – левая).
2. Сопоставить мерку с данными размерной таблицы.
Дело в том, что доминирующая рука больше подвергается механическим и физическим нагрузкам, поэтому, как правило, она более широкая и развитая. Если ориентироваться на противоположную руку, то изделие может быть мало. При необходимости можно измерить обе кисти. Если мерки не совпадают, то ориентиром должен быть наибольший обхват. Этот метод поможет в том случае, если ребенок телосложением отличается от сверстников.
Если ориентироваться на противоположную руку, то изделие может быть мало. При необходимости можно измерить обе кисти. Если мерки не совпадают, то ориентиром должен быть наибольший обхват. Этот метод поможет в том случае, если ребенок телосложением отличается от сверстников.
Выбор детских перчаток по возрасту
Если демисезонный или зимний аксессуар приобретается в подарок, то определить его ориентировочный размер можно по возрасту ребенка. В этом поможет специальная таблица:
|
Возраст ребенка |
Международный размер |
Российский размер |
|
0-6 месяцев |
- |
10 |
|
6-12 месяцев |
0 |
11 |
|
1-2 года |
1 |
12 |
|
2-3 года |
2 |
13 |
|
4-6 лет |
3 |
14 |
|
7-8 лет |
4 |
15 |
|
9-10 лет |
5 |
16 |
|
11-12 лет |
6 |
17 |
|
13-14 лет |
6 |
18 |
|
15-16 лет |
7 |
19 |
|
17 лет |
8 |
20 |
Таблица дана только в качестве ориентира.![]() На деле может оказаться, что у подростка узкая кисть, а у малыша – широкое запястье и длинные пальцы.
На деле может оказаться, что у подростка узкая кисть, а у малыша – широкое запястье и длинные пальцы.
Покупая перчатки, учитывают индивидуальные анатомические особенности конкретного ребенка. Если есть возможность, с этой целью можно использовать старую пару и, ориентируясь на нее, взять модель на 1-2 размера больше.
Полезные советы
Размер – не единственный параметр выбора детских перчаток и варежек. Помимо него нужно учитывать:
1. Материалы. Предпочтительнее натуральная шерсть или ангора с невысоким содержанием синтетических волокон. Так изделие будет хорошо сидеть на руке, дарить приятные тактильные ощущения, сохранять форму после стирок или длительной носки.
2. Качество вязки. Для температуры воздуха выше -10 градусов подойдут однослойные модели. В условиях суровой зимы не обойтись без перчаток двойной вязки из мериносовой или ангорской шерсти. Синтетические нити не должны составлять более 10-15 % состава.
3. Резинка. Чтобы изделие надежно держалось на руке, у него должна быть мягкая эластичная резинка. Предпочтительнее модели с длинной манжетой, которые хорошо защищают запястье. Резинка не должна быть слишком тугой, иначе будет нарушаться кровообращение.
4. Влагостойкость. Зима – время, предназначенное для игр в снежки, строительства ледяных крепостей и катаний с горок. Чтобы руки малыша не мерзли, пока он играет со снегом, у перчаток должно быть влагостойкое покрытие. Некоторые модели изготавливаются из материалов вроде плащевки или мембраны. Они защищают руки от влаги и ветра.
5. Декор. Зимний аксессуар может быть выдержан в яркой расцветке, украшен необычными вставками, интересным принтом, аппликацией, бусинами или стразами.
Как и другая детская одежда, варежки и перчатки должны нравиться ребенку. Если аксессуар подобран по размеру и соответствуют предпочтениям малыша, он обеспечит свободу движений и согреет даже в лютый мороз.
Дактилоскопия по фото ладони доказала причастность бобруйчанина к незаконному обороту наркотиков
Фото ГКСЭ
23 августа, Могилев /Корр. БЕЛТА/. Хранившиеся на смартфоне фото жителя Бобруйска помогли доказать его причастность к незаконному обороту наркотиков. Об этом БЕЛТА сообщила главный специалист по работе со СМИ управления Государственного комитета судебных экспертиз по Могилевской области Ирина Рачковская.
В Бобруйске недавно осудили 25-летнего местного жителя. «Правоохранители уличили парня в незаконном обороте наркотических средств — хранении марихуаны. У следствия имелись предположения, что парень лично выращивал коноплю для изготовления марихуаны, но доказательств не было. В поиске улик помог смартфон обвиняемого», — рассказала Ирина Рачковская.
Во время расследования у молодого человека изъяли мобильный телефон, в памяти которого среди прочего были обнаружены две фотографии, привлекшие особое внимание следствия. На фотографиях крупным планом изображена ладонь человека с растением, внешне схожим с коноплей. «Требовалось установить, чья же это рука. Следователь предоставил судебным экспертам Бобруйского межрайонного отдела Государственного комитета судебных экспертиз на исследование эти фотографии и дактилоскопическую карту с отпечатками рук обвиняемого. Отличное качество фотографий позволило судебным экспертам без каких-либо сложностей провести исследование папиллярного узора ладони, сфотографированной на камеру телефона», — рассказала главный специалист.
По результатам экспертизы установлены совпадения по общим признакам, а также по наличию и расположению деталей строения папиллярных узоров. Судебная дактилоскопическая экспертиза доказала, что изображения ладони на фотографиях принадлежат левой руке обвиняемого.
Заключение эксперта стало одним из доказательств причастности 25-летнего бобруйчанина к незаконному обороту наркотических средств. Под тяжестью собранных улик, в том числе результатов дактилоскопической экспертизы, обвиняемый признал вину в совершенном преступлении. Уголовное дело рассмотрено судом, молодому человеку назначено наказание в виде трех с половиной лет лишения свободы в исправительной колонии в условиях общего режима. Приговор вступил в законную силу.
Как подчеркнула Ирина Рачковская, это была первая дактилоскопическая экспертиза по фотографиям ладоней в практике судебных экспертов УГКСЭ по Могилевской области.-0-
Сугробы со вкусом арбуза и снежинки размером с ладонь: удивительные факты о снеге
Сколько на самом деле лучей у снежинки и как ее строение вдохновляет математиков и создателей игр? Где выпадает больше всего снега? И почему он бывает не только белый? Публикуем краткий пересказ ток-шоу, которое состоялось на площадке ИЦАЭ.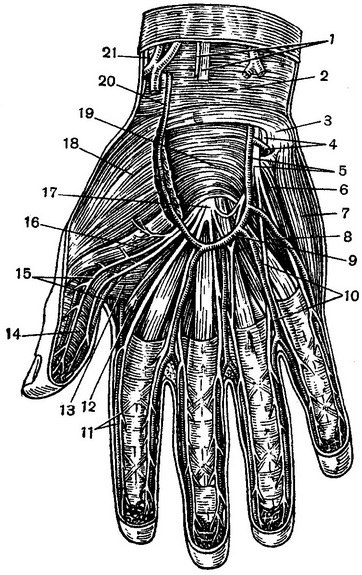 Мероприятие прошло в рамках проекта «Разберем на атомы».
Мероприятие прошло в рамках проекта «Разберем на атомы».
В ходе ток-шоу ученые, которые представляют различные отрасли знаний, «разбирают на атомы» общую тему. На этот раз математик и метеоролог посмотрели с необычных точек зрения на снег.
Профессор Валентина Горбатенко, заведующая кафедрой метеорологии и климатологии Томского государственного университета, рассказала о том, почему бывает разноцветный снег, от чего зависит форма снежинок и почему в пустыне Сахара все чаще идет снегопад.
А Михаил Семёнов, доцент Отделения экспериментальной физики Инженерной школы ядерных технологий Томского политехнического университета, рассказал о математических определениях, которые позволяют описывать форму снежинок, о том, что такое снежинка Коха и как эта природная форма вдохновляет компьютерщиков.
Источник: shutterstock.com
Для нас, как жителей одной из самых заснеженных стран, снег является вполне обычным явлением, которое мы наблюдаем каждый год. Но на самом деле половина всего населения Земли никогда не видели снега вживую. Впрочем, из-за процессов глобального потепления снегопады все чаще случаются в совсем неожиданных местах, например, в пустыне Сахара за последние 40 лет это случалось уже трижды. 7 января 2018 года там выпал 40 сантиметровый слой снега, который пролежал довольно долго и растаял только к вечеру.
Но на самом деле половина всего населения Земли никогда не видели снега вживую. Впрочем, из-за процессов глобального потепления снегопады все чаще случаются в совсем неожиданных местах, например, в пустыне Сахара за последние 40 лет это случалось уже трижды. 7 января 2018 года там выпал 40 сантиметровый слой снега, который пролежал довольно долго и растаял только к вечеру.
Метеорологи связывают это с процессом, называемым меридиональной циркуляцией: из-за происходящих в последние десятилетия изменений климата теплые воздушные массы выносит все дальше и глубже в Арктику, а холодные массы оттуда ― все дальше на юг.
При этом чаще всего снег выпадает не в северных регионах земли и не в Арктике, а в городе Аомори на севере Японии. В Японии же находится и единственный в мире музей снежинок, названный в честь основателя Накайя Укисиро ― специалиста по физике атмосферного льда, получившего известность благодаря исследованию природных ледяных кристаллов.
Кому-то может показаться удивительным, но к самым холодным странам мира относятся Казахстан и Монголия ― там средняя температура равняется нулю градусов Цельсия.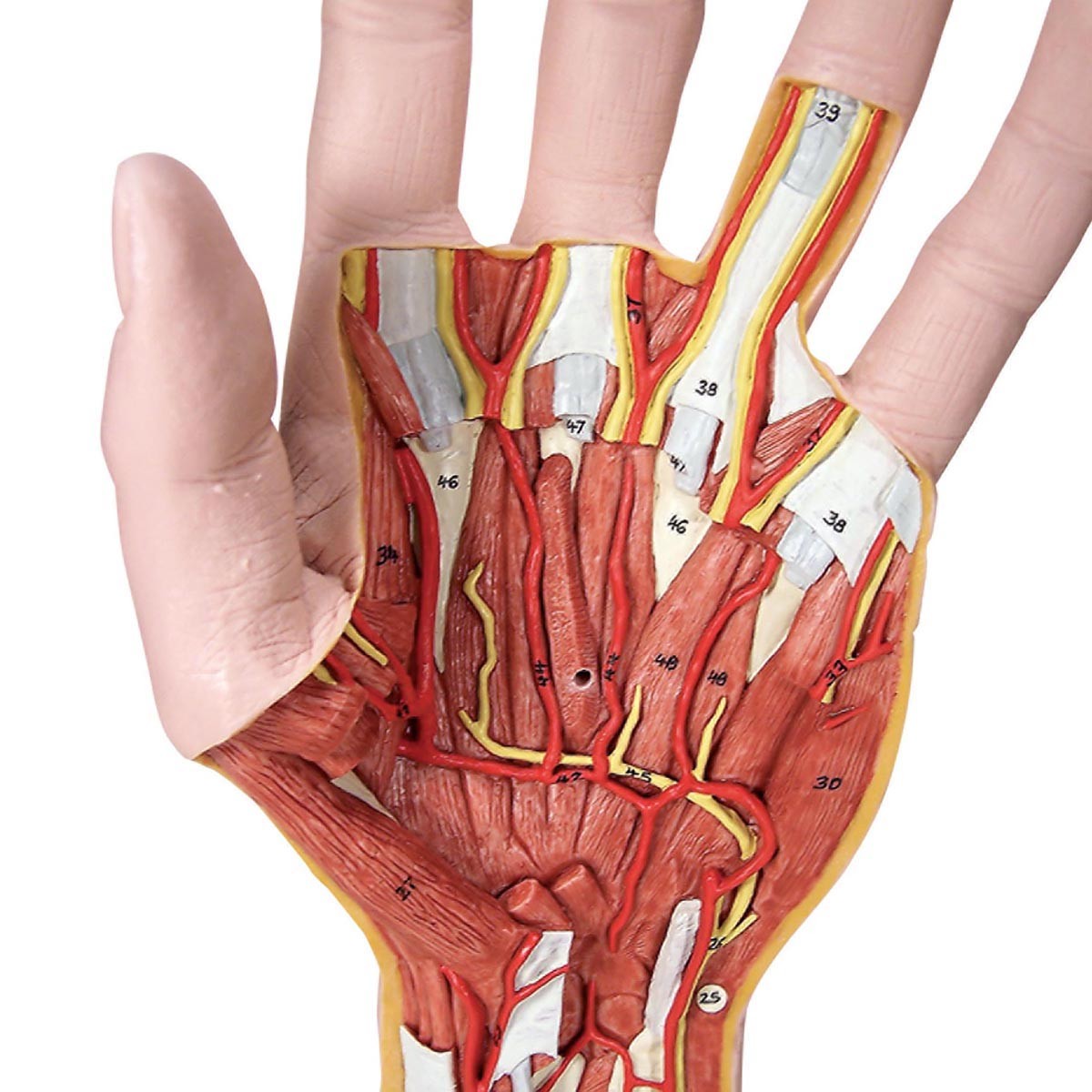 Но, конечно, наибольшее количество снега лежит в Антарктиде: снежное покрытие там может достигать 3 километров в высоту.
Но, конечно, наибольшее количество снега лежит в Антарктиде: снежное покрытие там может достигать 3 километров в высоту.
Музей снежинок Накайя Укисиро. Источник: allkaga.com
Одним из самых удивительных явлений на планете можно назвать выпадение цветного снега ― впрочем, это случается чаще, чем вы думаете. Например, желтый снег могли наблюдать жители Омской области в 2007 году, а розовый ― жители Приморского края в 2006-м. В 2002 году разноцветный снег выпал на Камчатке, а в 2006 году жители Колорадо столкнулись с удивительным «арбузным снегом», который не только обладал характерным ярким цветом, но имел запах и даже вкус арбуза.
Снег бывает также зеленым и даже фиолетовым. Виной этому ― особые водоросли под названием хламидомонада снежная, которые живут только в холодных условиях и погибают при -4 градусах.
Во многих культурах белый цвет считается символом смерти и зла — и это может понять любой, кто оказался на Крайнем Севере во время полярной ночи. У норвежцев есть пословица: «Северный ветер создал викингов» — суровых, мужественных, аскетичных, но в то же время крайне отзывчивых и сильных людей.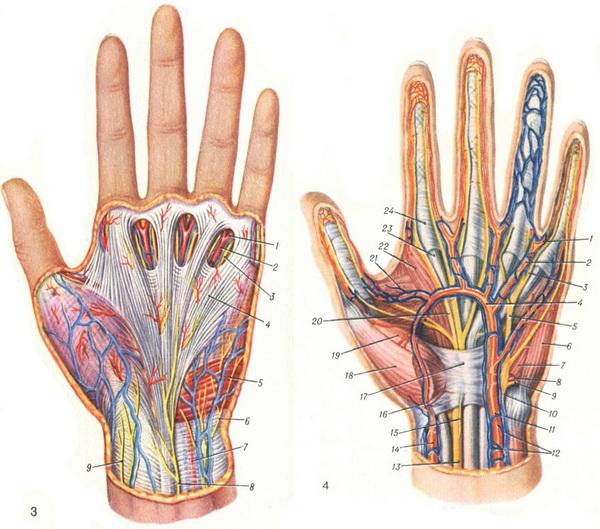 Особый скандинавский менталитет, который многие описывают как флегматичный, специалисты считают прямым следствием условий их жизни.
Особый скандинавский менталитет, который многие описывают как флегматичный, специалисты считают прямым следствием условий их жизни.
Розовый снег на склонах в Калифорнии. Источник: wikipedia.org
Раньше считалось, что больше всего слов, связанных с обозначением различных видов снега и льда, у эскимосов. Но их победили саамы, живущие на севере Скандинавии, — в их лексиконе насчитывается как минимум 180 таких слов. Например, свои обозначения есть у первого и последнего снегопада в году, а также вчерашнего и завтрашнего; у снега, подходящего и не подходящего для строительства, у медленно или быстро идущего снегопада. Отдельным словом называется снег, заметающий всю деревню.
То, что снег ― это просто замерзшие капли дождя, большое заблуждение. На самом деле влага из воздуха из газообразного состояния сразу превращается в твердые кристаллики льда, минуя стадию жидкости. Такой процесс «перескакивания» через агрегатное состояние называется сублимацией.
В среднем снежинки имеют диаметр 5 мм и вес около 0,004 гр. Но бывают и исключения, например, 30 апреля 1944 года на территории Москвы видели снежинки размером с человеческую ладонь. Особую шестиконечную форму снежинок впервые описал шведский пастор Олаус Магнус в 1555 году. Он же был первым, кто заметил, что каким бы замысловатым ни был узор, мотив шестиугольника всегда повторяется.
Но бывают и исключения, например, 30 апреля 1944 года на территории Москвы видели снежинки размером с человеческую ладонь. Особую шестиконечную форму снежинок впервые описал шведский пастор Олаус Магнус в 1555 году. Он же был первым, кто заметил, что каким бы замысловатым ни был узор, мотив шестиугольника всегда повторяется.
Формы снежинок. Иллюстрация из лекции
Впрочем, снежинки далеко не всегда имеют красивую форму дендритов ― именно так называется классическая в нашем представлении звездчатая форма. Снежинки бывают в виде пластин, стобиков, полых столбиков, колонн, игл и так далее. Это прямым образом зависит от условий окружающей среды: сочетаний показателей температуры и влажности воздуха в различных комбинациях.
Например, звезды получаются при -2 и -15, а вот при -5 или -10 формируются колонны и стройные иглы. А еще на форму снежинок влияет ветер: чем он слабее, тем больше их размер, а вот сильный ветер ломает их и превращает в «осколки». Подробную схему зависимости температурных условий и влажности на форму снежинок описал тот самый Накайя Укисиро, который основал музей в Японии.
Источник: shutterstock.com
Шестикратная симметрия возникает из-за расположения молекул воды в решетке кристаллического льда. При вращении модели h3O можно отчетливо увидеть шестиугольники. Простая формула, по которой строятся снежинки, описывается в теории нечетких автоматов и активно используется в компьютерном моделировании. А сама схема снежинки служит вдохновением для проектирования хранилищ и баз данных.
Снежинка Коха ― одна из самых известных фрактальных кривых. Она строится из равностороннего простого треугольника: каждая сторона делится на три равные части и вмещает в центральной части такой же равносторонний треугольник. Если убрать основания треугольников, как раз получается шестиконечный дендрит ― снежинка. Далее процесс итеративно повторяется.
Вы можете сами попробовать построить фрактальные снежинки, используя онлайн-сервисы, например, по ссылке. Это простейшая демонстрация фрактальной графики, которая используется, например, в создании компьютерных игр для генерации карт и ландшафтов.
Полную версию лекции можно посмотреть на канале сети ИЦАЭ в YouTube.
Перейти к содержаниюENh2212/EP473: Морфология и анатомия ладони
Пальмы сильно отличаются от широколиственных (двудольных) и хвойных (голосеменных) деревьев как по общей форме и внешнему строению (морфология), так и по внутреннему строению (анатомия). Морфология и анатомия определяют, как ладони растут, функционируют и реагируют на внешние и внутренние факторы стресса. Эта публикация дает общее представление о том, как устроены пальмы.
Стебли
Стебли пальм (стволы) значительно различаются по размерам и внешнему виду у разных видов, но в целом они имеют цилиндрическую или слегка сужающуюся форму, а иногда и выпуклую форму (рис. 1).Поверхность может быть от гладкой до чрезвычайно шероховатой и узловатой, а также может быть вооружена острыми шипами. У некоторых видов на стеблях видны заметные и часто привлекательные шрамы от листьев, которые были точками прикрепления листьев к стеблю. Расстояние между этими листовыми рубцами (междоузлиями) зависит от скорости роста стебля. У молодых пальм образуются самые длинные междоузлия, длина которых резко уменьшается по мере взросления пальмы.
У некоторых видов на стеблях видны заметные и часто привлекательные шрамы от листьев, которые были точками прикрепления листьев к стеблю. Расстояние между этими листовыми рубцами (междоузлиями) зависит от скорости роста стебля. У молодых пальм образуются самые длинные междоузлия, длина которых резко уменьшается по мере взросления пальмы.
Кредит: А.В. Мироу, UF/IFAS
Стебли молодых пальм (тех, которые еще не достигли своего максимального диаметра) удлиняются очень медленно, постепенно увеличиваясь в диаметре с каждым новым листом. В результате основания стебля пальмы имеют V-образную форму, причем вершина буквы V соответствует самой ранней ткани стебля сеянца (рис. 2). Как только ювенильный стебель пальмы достигает максимального диаметра, начинается существенное вертикальное удлинение стебля, и дальнейшее увеличение диаметра стебля практически прекращается.Набухание, наблюдаемое у основания стебля у зрелых экземпляров некоторых видов, обычно является результатом развития новых корней внутри стебля, хотя этому набуханию также может способствовать увеличение клеток паренхимы или отложение лигнина.
Авторы и права: Т. К. Брошат, UF/IFAS
Пальмы могут быть одноствольными или многоствольными (группировками) в результате ответвления от пазушных почек, расположенных низко на стебле.Только несколько видов (например, Hyphaene spp.) обычно демонстрируют воздушное ветвление.
Стебли пальмы характеризуются наличием единственной апикальной меристемы или точки роста, которую также называют почкой или сердцевиной. Все новые листья и цветки развиваются из апикальной меристемы (рис. 3). Он окружен основаниями листьев, а у видов с венчиком расположен вблизи границы венчика и стебля. У пальм нет боковых меристем или сосудистого камбия, которые у двудольных и хвойных деревьев образуют дополнительную ксилему внутри и флоэму и кору снаружи.Смерть апикальной меристемы на пальме приводит к гибели этого стебля у кластерных видов и гибели всей пальмы у одностебельных видов. Это также означает, что стволовые раны, которые в конечном итоге будут разделены и зарастут у двудольных деревьев, на пальмах остаются постоянными.
Это также означает, что стволовые раны, которые в конечном итоге будут разделены и зарастут у двудольных деревьев, на пальмах остаются постоянными.
Кредит: Т.К. Брошат, UF/IFAS
Анатомически стебли пальм типичны для однодольных, с ксилемой (водопроводящая ткань) и флоэмой (углеводопроводящая ткань), примыкающими к сосудистым пучкам, разбросанным по всему центральному цилиндру стебля. У большинства видов эти пучки сосредоточены вблизи периферии стебля (рис. 4). Они вкраплены в матрицу тонкостенных недифференцированных клеток паренхимы. Tomlinson (1990) описывает стволы пальм как аналоги железобетонных столбов, с сосудистыми пучками, эквивалентными стальным стержням, и клетками паренхимы, аналогичными бетону.Волокнистые клетки, прилегающие к флоэме внутри сосудистых пучков, продолжают откладывать лигнин и целлюлозу на протяжении всей своей жизни, тем самым укрепляя самые старые части стебля пальмы. В стеблях двудольных и хвойных деревьев клетки сосудов ксилемы отмирают и теряют свое содержимое до того, как становятся функционирующими в качестве водопроводной ткани, и постоянно образуется новая флоэма, заменяющая старую. Однако клетки ксилемы, флоэмы и даже паренхимы ствола пальмы остаются живыми на протяжении всей жизни пальмы, которая у некоторых видов может составлять сотни лет (Tomlinson and Huggett 2012).Снаружи от центрального цилиндра находится область склерозированной ткани, известная как кора, и очень тонкий эпидермис, которые иногда вместе называют «псевдобаркой».
В стеблях двудольных и хвойных деревьев клетки сосудов ксилемы отмирают и теряют свое содержимое до того, как становятся функционирующими в качестве водопроводной ткани, и постоянно образуется новая флоэма, заменяющая старую. Однако клетки ксилемы, флоэмы и даже паренхимы ствола пальмы остаются живыми на протяжении всей жизни пальмы, которая у некоторых видов может составлять сотни лет (Tomlinson and Huggett 2012).Снаружи от центрального цилиндра находится область склерозированной ткани, известная как кора, и очень тонкий эпидермис, которые иногда вместе называют «псевдобаркой».
Авторы и права: Т. К. Брошат, UF/IFAS
Корни
Корни пальм сильно отличаются от корней широколиственных и хвойных деревьев тем, что все они являются придаточными, выходящими из области ствола, называемой зоной заложения корней. Корни пальмы берут начало из внешней области центрального цилиндра, где они соединяются с сосудистыми пучками стебля. По мере их роста огромный объем новых корней может привести к расщеплению коры и псевдокоры и расширению их от основания стебля (рис. 5). Если новые корни развиваются над землей и вступают в контакт с сухим воздухом, их развитие будет остановлено до тех пор, пока не возникнут более благоприятные условия. Если почву или мульчу насыпать вокруг зачатков воздушных корней, они возобновят свой рост вглубь почвы.Корни пальм вырастают в своем максимальном диаметре, и дополнительного увеличения диаметра не происходит. Некоторые первичные корни могут расти вниз, но большинство растут в стороны, у крупных экземпляров они отходят от ствола на 50 футов и более. Если первичные корни срезать, они могут разветвляться из-за среза, но полученные корни будут первичными по размеру и морфологии. Корни вторичного, третичного и даже четвертого порядка пропорционально меньше в диаметре и длине, недолговечны и часто растут вверх к поверхности почвы.
Корни пальмы берут начало из внешней области центрального цилиндра, где они соединяются с сосудистыми пучками стебля. По мере их роста огромный объем новых корней может привести к расщеплению коры и псевдокоры и расширению их от основания стебля (рис. 5). Если новые корни развиваются над землей и вступают в контакт с сухим воздухом, их развитие будет остановлено до тех пор, пока не возникнут более благоприятные условия. Если почву или мульчу насыпать вокруг зачатков воздушных корней, они возобновят свой рост вглубь почвы.Корни пальм вырастают в своем максимальном диаметре, и дополнительного увеличения диаметра не происходит. Некоторые первичные корни могут расти вниз, но большинство растут в стороны, у крупных экземпляров они отходят от ствола на 50 футов и более. Если первичные корни срезать, они могут разветвляться из-за среза, но полученные корни будут первичными по размеру и морфологии. Корни вторичного, третичного и даже четвертого порядка пропорционально меньше в диаметре и длине, недолговечны и часто растут вверх к поверхности почвы. Это корни, которые поглощают воду и питательные вещества. Специализированные корни, называемые пневматофорами, могут буквально расти вверх и из земли у некоторых видов (например, Phoenix spp.). Корни пальм не имеют корневых волосков, как корни двудольных. Поскольку у них нет камбия, корни соседних пальм не могут привиться, как у двудольных деревьев.
Это корни, которые поглощают воду и питательные вещества. Специализированные корни, называемые пневматофорами, могут буквально расти вверх и из земли у некоторых видов (например, Phoenix spp.). Корни пальм не имеют корневых волосков, как корни двудольных. Поскольку у них нет камбия, корни соседних пальм не могут привиться, как у двудольных деревьев.
Авторы и права: Т. К. Брошат, UF/IFAS
Листья
Область пальмы, на которой расположены листья, называется кроной или кроной. Пальмовые листья обычно бывают трех типов. Перистые или перистые листья имеют листочки, полностью отделенные друг от друга и прикрепленные перпендикулярно к оси, продолжению черешка в листовую пластинку (рис. 6). Примером перистолистной пальмы является кокосовая пальма. У пальчатолистных пальм (веерных пальм) соседние листочки или листовые сегменты соединены латерально на некоторой или большей части их длины.Они происходят из одной точки на кончике черешка, который часто включает в себя специальный выступ, называемый хастулой. Пальмы Washingtonia — типичные пальмовидные пальмы. Костапальчатые листья занимают промежуточное положение между перистыми и пальчатыми листьями, при этом общая листовая пластинка имеет форму от круглой до овальной. Листочки соединены вместе на некоторой или большей части своей длины, но прикрепляются вдоль косты, которая является продолжением черешка в листовую пластинку. видов Sabal имеют реберно-пальчатые листья.Некоторые другие формы листьев также могут встречаться среди пальм. Некоторые видов Chamaedorea имеют раздвоенные или двухлопастные листья, а видов Caryota имеют двуперистые листья. Пальмовые листочки обычно имеют V-образную форму в поперечном сечении со средней жилкой на вершине V.
У пальчатолистных пальм (веерных пальм) соседние листочки или листовые сегменты соединены латерально на некоторой или большей части их длины.Они происходят из одной точки на кончике черешка, который часто включает в себя специальный выступ, называемый хастулой. Пальмы Washingtonia — типичные пальмовидные пальмы. Костапальчатые листья занимают промежуточное положение между перистыми и пальчатыми листьями, при этом общая листовая пластинка имеет форму от круглой до овальной. Листочки соединены вместе на некоторой или большей части своей длины, но прикрепляются вдоль косты, которая является продолжением черешка в листовую пластинку. видов Sabal имеют реберно-пальчатые листья.Некоторые другие формы листьев также могут встречаться среди пальм. Некоторые видов Chamaedorea имеют раздвоенные или двухлопастные листья, а видов Caryota имеют двуперистые листья. Пальмовые листочки обычно имеют V-образную форму в поперечном сечении со средней жилкой на вершине V. Пальмовые листья с вертикальными V-образными листочками называются удвоенными, а листочки с перевернутой V-образной формой называются повторными.
Пальмовые листья с вертикальными V-образными листочками называются удвоенными, а листочки с перевернутой V-образной формой называются повторными.
Кредит: А.В. Мироу, UF/IFAS
Листья пальмы
обычно имеют черешки разной длины, но у некоторых видов Copernicia черешок отсутствует. Все пальмовые листья прикреплены к стволу расклешенным основанием. У некоторых пальм (например, Roystonea spp. , Dypsis spp. и т. д.) основания листьев имеют трубчатую форму и обвиваются друг вокруг друга, образуя гладкую стеблеобразную структуру, называемую коронным валом.
Цветы
Цветоносы (соцветия) возникают в пазухах листьев у большинства видов пальм, но у тех видов, у которых есть стебель, соцветия выходят из стебля чуть ниже основания стебля (рис. 7).У некоторых гапаксантических (пальмы, которые отмирают после цветения) видов, таких как Arenga и Caryota , соцветия развиваются из пазушных почек по всей длине стебля после перехода пальмы из вегетативного состояния в репродуктивное. У других видов гапаксантических пальм, таких как Corypha или Metroxylon , соцветие верхушечное, расположенное над кроной (рис. 8). Развивающиеся соцветия обычно покрыты защитным прицветником, называемым профиллом.Первичный стебель соцветия известен как цветонос, а ветви первого и второго порядка называются осью и рахиллой соответственно. На однодомных пальмах встречаются как мужские, так и женские цветки, в то время как на двудомных пальмах есть только мужские или женские цветки.
У других видов гапаксантических пальм, таких как Corypha или Metroxylon , соцветие верхушечное, расположенное над кроной (рис. 8). Развивающиеся соцветия обычно покрыты защитным прицветником, называемым профиллом.Первичный стебель соцветия известен как цветонос, а ветви первого и второго порядка называются осью и рахиллой соответственно. На однодомных пальмах встречаются как мужские, так и женские цветки, в то время как на двудомных пальмах есть только мужские или женские цветки.
Авторы и права: Т. К. Брошат, UF/IFAS
Рисунок 8. Верхнее соцветие у Corypha umbraculifera , гапаксановой пальмы.
Авторы и права: Т. К. Брошат, UF/IFAS
Плоды пальмы ботанически классифицируются как костянки и имеют одно или несколько костистых семян, покрытых мясистым, мучнистым или волокнистым мезокарпием и тонким поверхностным эпикарпием. Мезокарпий некоторых видов (например, Caryota, Arenga, Roystonea, Chamaedorea и др.) содержит игольчатые кристаллы оксалата кальция, которые сильно раздражают открытые участки кожи. Каменистый внешний слой семени известен как эндокарпий, который заключает в себе ткань для хранения пищи (эндосперм) и зародыш.
Мезокарпий некоторых видов (например, Caryota, Arenga, Roystonea, Chamaedorea и др.) содержит игольчатые кристаллы оксалата кальция, которые сильно раздражают открытые участки кожи. Каменистый внешний слой семени известен как эндокарпий, который заключает в себе ткань для хранения пищи (эндосперм) и зародыш.
Ссылки
Tomlinson, P.B. 1990. Структурная биология пальм. Оксфорд, Великобритания: Clarendon Press.
Томлинсон, П. Б. и Б. А. Хаггетт. 2012. «Продолжительность жизни клеток и устойчивый первичный рост стеблей пальм». амер. Дж. Бот. 99: 1891–1902.
Пальмы — Структура — Ствол, Дерево, Ширина и Листья
Пальмы в основном представляют собой неразветвленные кустарники или деревья и являются основным семейством деревьев в составе однодольных растений.Обычно одиночный прямостоячий стебель увенчан крупными устойчивыми листьями, покрывающими основание. Листья Raphia fainifera являются самыми большими из всех цветковых растений , иногда достигая более 65 футов (20 м). Пальмовые листья иногда бывают простыми, но обычно они рассечены на веерообразные (пальчатые) или перистые со множеством отчетливых сегментов, которые проходят перпендикулярно главной оси листа (перистосложные). Стебель может быть очень коротким, чтобы казаться практически отсутствующим, до 164 футов (50 м) в высоту.Не все пальмы имеют одиночные стебли или стволы. Некоторые пальмы имеют сгруппированные стебли, которые возникают из почек у основания исходного стебля, например, Феникс. У некоторых необычных видов, таких как Chrysalidocarpus madagascariensis , одни особи имеют одиночные прямые стволы, а другие — со сгруппированными стволами. Некоторые пальмы лазают по лозам, например, ротанговые.
Пальмовые листья иногда бывают простыми, но обычно они рассечены на веерообразные (пальчатые) или перистые со множеством отчетливых сегментов, которые проходят перпендикулярно главной оси листа (перистосложные). Стебель может быть очень коротким, чтобы казаться практически отсутствующим, до 164 футов (50 м) в высоту.Не все пальмы имеют одиночные стебли или стволы. Некоторые пальмы имеют сгруппированные стебли, которые возникают из почек у основания исходного стебля, например, Феникс. У некоторых необычных видов, таких как Chrysalidocarpus madagascariensis , одни особи имеют одиночные прямые стволы, а другие — со сгруппированными стволами. Некоторые пальмы лазают по лозам, например, ротанговые.
Ствол пальм сильно отличается от ствола хвойных или двудольных деревьев, преобладающих в регионах с умеренным климатом.Хвойные деревья, такие как сосны , ели и болиголовы, а также двудольные деревья, такие как клены , дубы и вязы, увеличивают ширину своего ствола по мере роста за счет процесса, называемого вторичным ростом.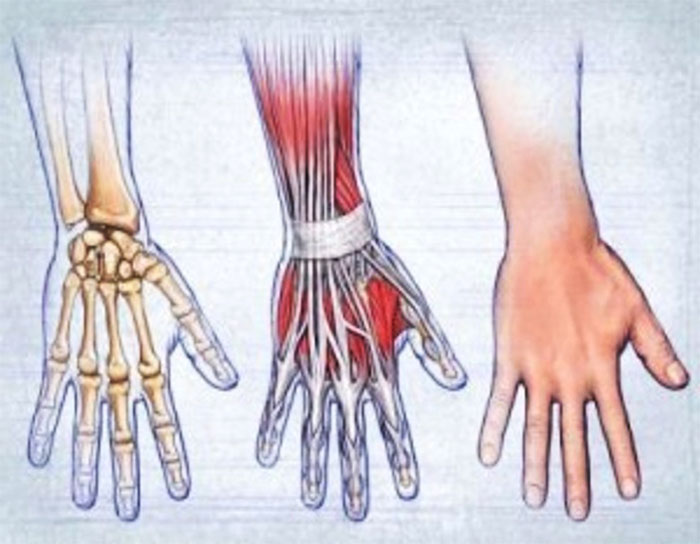 Во время этого процесса кольцо специализированных клеток под корой дерева производит новую древесину по направлению к центру дерева и другие виды специализированных проводящих тканей по направлению к стороне коры. Вторичный рост на ладонях отсутствует. Вместо этого, когда семя прорастает, росток сначала превращается в перевернутый конус, ширина которого соответствует полной ширине выращиваемого ствола.Только после завершения этого радиального роста сеянец начинает расти вертикально, сохраняя свою ширину. Если гвоздь вбить в ствол хвойного или двудольного дерева, дерево вырастет вокруг него, в конечном итоге полностью погрузив гвоздь в ствол. Гвоздь, вбитый в ствол пальмы, останется там, где вбит, и не застрянет в стволе.
Во время этого процесса кольцо специализированных клеток под корой дерева производит новую древесину по направлению к центру дерева и другие виды специализированных проводящих тканей по направлению к стороне коры. Вторичный рост на ладонях отсутствует. Вместо этого, когда семя прорастает, росток сначала превращается в перевернутый конус, ширина которого соответствует полной ширине выращиваемого ствола.Только после завершения этого радиального роста сеянец начинает расти вертикально, сохраняя свою ширину. Если гвоздь вбить в ствол хвойного или двудольного дерева, дерево вырастет вокруг него, в конечном итоге полностью погрузив гвоздь в ствол. Гвоздь, вбитый в ствол пальмы, останется там, где вбит, и не застрянет в стволе.
Пальмовые цветки иногда обоеполые, но обычно однополые. У однополых цветки каждого пола могут быть на одном и том же растении или, как у человека, на особь приходится только один пол .Цветки маленькие и обычно располагаются на больших разветвленных стеблях (соцветиях), которые расположены внутри кроны или чуть ниже нее. Цветок Детали обычно состоят из трех частей. Биология опыления пальм изучена недостаточно, тем не менее в семействе распространены как ветровое опыление, так и опыление насекомыми.
Цветок Детали обычно состоят из трех частей. Биология опыления пальм изучена недостаточно, тем не менее в семействе распространены как ветровое опыление, так и опыление насекомыми.
Пальма Плоды — это ягоды, которые представляют собой простые мясистые плоды, содержащие одно или несколько семян , или костянки, которые представляют собой простые мясистые плоды, содержащие одно семя, окруженное костлявой ямкой, косточковый плод.Самый большой плод в мире происходит от двойной кокосовой пальмы ( Lodoicea sechellarum ) и весит до 40 фунтов (18,2 кг).
пальма | дерево | Британника
Распространение
Крупнейшие центры распространения пальм находятся в Америке и Азии от Индии до Японии и на юг до Австралии и островов Тихого и Индийского океанов, с Африкой и Мадагаскаром в качестве третьего, но гораздо менее важного региона выращивания пальм. Эти центры примерно ограничены 44° северной широты и 44° южной широты, хотя наибольшая численность находится в основном между 30° северной широты и 30° южной широты. Эти распределения более или менее соответствуют зонам со среднегодовыми температурами 15 и 21 ° C (60 и 70 ° F) и, за редким исключением, районам с 500 мм (20 дюймов) или более осадков в год.
Эти распределения более или менее соответствуют зонам со среднегодовыми температурами 15 и 21 ° C (60 и 70 ° F) и, за редким исключением, районам с 500 мм (20 дюймов) или более осадков в год.
Британская викторина
Деревья мира
Что такое ладан? Что такое лиственное дерево? Посадите свои корни в этой викторине и проверьте свои знания о деревьях мира.
Самая северная пальма — европейская веерная пальма ( Chamaerops humilis ), которая растет в Средиземноморье в Европе и Северной Африке; самая южная — пальма никау ( Rhopalostylis sapida ) Новой Зеландии и островов Чатем. Хотя есть виды с обширным ареалом, особенно в Америке, большинство из них имеет ограниченный ареал, и, в частности, островные виды часто больше нигде не встречаются. Один вид, Maxburretia gracilis , ограничен несколькими обнажениями известняка на островах Лангкави у Малайского полуострова. На острове Новая Каледония насчитывается 17 родов и 32 вида пальм, все они эндемичны. Пальмы Мадагаскара еще малоизвестны, но зарегистрировано 130 видов, принадлежащих к 21 роду, и не менее 14 родов встречаются только на острове. Ни один вид, за исключением европейской веерной пальмы и пантропического культивируемого кокоса ( Cocos nucifera ), не встречается более чем на одном континенте; роды, выходящие за континентальные границы, — это Chamaerops в Европе и Африке, Elaeis (масличная пальма) и Raphia (пальма рафия или юпати) в Африке и Америке и Borassus (пальмировая пальма), Calamus ( ротанговая пальма), Hyphaene (пальма дум) и Phoenix (финиковая пальма) в Африке и Азии.Численность особей вида может быть как малой, так и многочисленной.
На острове Новая Каледония насчитывается 17 родов и 32 вида пальм, все они эндемичны. Пальмы Мадагаскара еще малоизвестны, но зарегистрировано 130 видов, принадлежащих к 21 роду, и не менее 14 родов встречаются только на острове. Ни один вид, за исключением европейской веерной пальмы и пантропического культивируемого кокоса ( Cocos nucifera ), не встречается более чем на одном континенте; роды, выходящие за континентальные границы, — это Chamaerops в Европе и Африке, Elaeis (масличная пальма) и Raphia (пальма рафия или юпати) в Африке и Америке и Borassus (пальмировая пальма), Calamus ( ротанговая пальма), Hyphaene (пальма дум) и Phoenix (финиковая пальма) в Африке и Азии.Численность особей вида может быть как малой, так и многочисленной.
Очаги обилия видов встречаются в смешанных тропических и субтропических лесах как в американских, так и в азиатских тропиках. Низменные тропические леса Новой Гвинеи и Зондских островов, а также тропические леса Центральной и Южной Америки богаты видами пальм. В регионе Чоко на западе Колумбии и в некоторых частях острова Борнео растет невероятное количество пальм. В национальном парке Гунунг-Мулу, Малайзия, на площади тропических лесов площадью около 52 864 га обитает 111 видов, представляющих 20 родов.В западной части региона Антиокия на северо-западе Колумбии на площади 19 000 квадратных километров (7 300 квадратных миль) встречается 89 видов, представляющих 34 рода пальм. Примечательно, что пальмы не являются доминирующим элементом в этих лесах. В некоторых типах растительности в тропиках и субтропиках преобладают крупные насаждения отдельных видов. Восковая пальма карнаубская ( Copernicia alba ) встречается сплошными насаждениями площадью сотни квадратных километров в северо-восточной части парагвайского Бореала Чако и прилегающих Боливии и Бразилии, самые большие насаждения только в этом регионе содержат, возможно, 500 миллионов растений.
В регионе Чоко на западе Колумбии и в некоторых частях острова Борнео растет невероятное количество пальм. В национальном парке Гунунг-Мулу, Малайзия, на площади тропических лесов площадью около 52 864 га обитает 111 видов, представляющих 20 родов.В западной части региона Антиокия на северо-западе Колумбии на площади 19 000 квадратных километров (7 300 квадратных миль) встречается 89 видов, представляющих 34 рода пальм. Примечательно, что пальмы не являются доминирующим элементом в этих лесах. В некоторых типах растительности в тропиках и субтропиках преобладают крупные насаждения отдельных видов. Восковая пальма карнаубская ( Copernicia alba ) встречается сплошными насаждениями площадью сотни квадратных километров в северо-восточной части парагвайского Бореала Чако и прилегающих Боливии и Бразилии, самые большие насаждения только в этом регионе содержат, возможно, 500 миллионов растений.
В восточных тропиках Nypa fruticans могут образовывать плотные колонии на эстуарных илах; эти чистые насаждения пальмы нипа ( Nypa ) простираются на сотни гектаров в восточной части Суматры и некоторых частях Борнео. В других ситуациях двудольные виды мангровых зарослей встречаются с пальмой нипа. Род Manicaria (пальма буссу) занимает аналогичные местообитания в некоторых районах Нового Света. Пальмы доминируют в другом типе растительности на береговой окраине мангровых болот в западной части Малайского архипелага, где встречаются Oncosperma tigillarium и Calamus erinaceus (и на Борнео Daemonorops longispathus ).В эстуарии Амазонки Raphia taedigera занимает обширные площади; другие виды пальмы рафия доминируют в аналогичных местах обитания в Западной Африке. Пальма рафия встречается в почти чистых насаждениях между болотами и двудольными заболоченными лесами вдоль Карибского и Тихоокеанского побережья Коста-Рики, а Mauritia flexuosa встречается на обширных насаждениях во внутренних частях бассейна Амазонки. На приречных отмелях и прибрежных равнинах Африки часто в большом количестве встречаются Hyphaene Compressa и Borassus aethiopum .
В других ситуациях двудольные виды мангровых зарослей встречаются с пальмой нипа. Род Manicaria (пальма буссу) занимает аналогичные местообитания в некоторых районах Нового Света. Пальмы доминируют в другом типе растительности на береговой окраине мангровых болот в западной части Малайского архипелага, где встречаются Oncosperma tigillarium и Calamus erinaceus (и на Борнео Daemonorops longispathus ).В эстуарии Амазонки Raphia taedigera занимает обширные площади; другие виды пальмы рафия доминируют в аналогичных местах обитания в Западной Африке. Пальма рафия встречается в почти чистых насаждениях между болотами и двудольными заболоченными лесами вдоль Карибского и Тихоокеанского побережья Коста-Рики, а Mauritia flexuosa встречается на обширных насаждениях во внутренних частях бассейна Амазонки. На приречных отмелях и прибрежных равнинах Африки часто в большом количестве встречаются Hyphaene Compressa и Borassus aethiopum . В пресноводных болотах в некоторых частях Новой Гвинеи преобладает вид Metroxylon sagu . И пальма дум, и пальма саговая ( Metroxylon ) полезны, и их распространение может быть частично связано с деятельностью человека. Eugeissona utilis растет в густых местных насаждениях, исключая другие деревья на возвышенностях Борнео. Растительность, в которой преобладает Prestoea montana , характерна для горных лесов Карибского бассейна. Многие из этих пальм экономически полезны, а их естественные или полуестественные насаждения могут иметь огромное значение для местной экономики.
В пресноводных болотах в некоторых частях Новой Гвинеи преобладает вид Metroxylon sagu . И пальма дум, и пальма саговая ( Metroxylon ) полезны, и их распространение может быть частично связано с деятельностью человека. Eugeissona utilis растет в густых местных насаждениях, исключая другие деревья на возвышенностях Борнео. Растительность, в которой преобладает Prestoea montana , характерна для горных лесов Карибского бассейна. Многие из этих пальм экономически полезны, а их естественные или полуестественные насаждения могут иметь огромное значение для местной экономики.
Численность пальм также может рассматриваться в связи с количеством видов на род, поскольку несколько родов пальм имеют большое количество видов. Calamus с примерно 379 — самый большой, а Bactris (персиковая пальма) с примерно 239 — второй. Несколько других родов, Licuala, Pinanga, Chamaedorea и Daemonorops , насчитывают более 100 видов каждый. Однако почти треть родов (64) имеют только один вид, а более половины имеют менее 5 видов каждый.Небольшое количество видов в роду отражает большой эндемизм в семействе.
Однако почти треть родов (64) имеют только один вид, а более половины имеют менее 5 видов каждый.Небольшое количество видов в роду отражает большой эндемизм в семействе.
Пальмовые листья | Ботанический мир приключений
Существует более 3000 различных видов пальм, но они, как правило, имеют лишь несколько основных узоров листьев. Двумя наиболее распространенными структурами листьев пальм являются веерообразные/пальчатые (, рис. 30 ) и перистые/перистые (, рис. 31, ).
Еще одна интересная характеристика пальм, связанная с листьями, заключается в том, являются ли они «самоочищающимися».Если весь лист чисто отделяется от ствола у основания листа и падает с дерева, то такая пальма называется самоочищающейся. Обычно по длине ствола таких пальм имеются «кольца», которые отмечают место прикрепления (узел) опавшего листа ( Рисунок 32 ).
Пальмовые листья могут быть очень большими и тяжелыми! Листья зрелых королевских пальм могут весить до 100 фунтов, а листовое влагалище может быть достаточно большим, чтобы обернуться вокруг человека, как хот-дог в булочке, в то время как листовая пластинка может достигать десяти футов в длину ( Рисунок 33 ). Они могут упасть без предупреждения, хотя чаще всего это происходит в ветреную погоду. Вы же не хотите оказаться под одним из них, когда он упадет!
Они могут упасть без предупреждения, хотя чаще всего это происходит в ветреную погоду. Вы же не хотите оказаться под одним из них, когда он упадет!
Посетители сада часто спрашивают, можно ли по кольцам на стволах пальм определить, сколько лет дереву. В самом общем смысле можно приблизительно определить возраст дерева, но далеко не так точно, как в случае с широколиственными деревьями из зон с умеренным климатом. Пальма производит внешние кольца ствола с той же скоростью, что и листья, а производство листьев напрямую связано с энергией дерева, которая, в свою очередь, связана с почвенными условиями (питательными веществами, водой и доступностью воздуха, а также климатическими факторами).Такую пальму пришлось бы изучать в течение нескольких лет и отслеживать ежегодное производство листьев. Затем можно было бы подсчитать общее количество колец вдоль ствола, разделить на среднегодовое количество листьев и получить приблизительный возраст дерева. Это, вероятно, не соответствует его истинному возрасту, потому что кольца, образованные листьями молодого сеянца, стираются расширением ствола у его основания.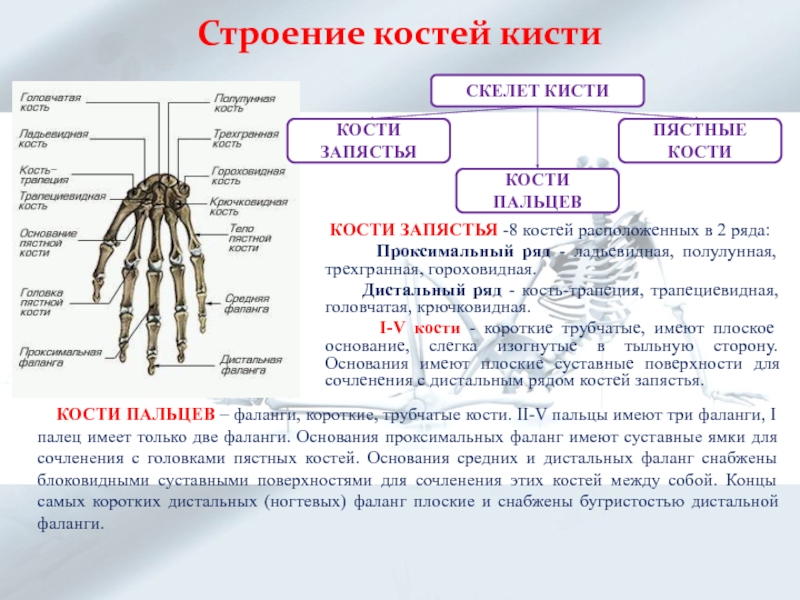
Однако у некоторых пальм лист просто отмирает и остается прикрепленным к стволу, часто в течение длительного времени, а затем может просто сгнить и отломиться где-нибудь у основания старого черешка или его придется отрезать пилой ( Фото 34 и 35 ).Эта ситуация может привести к увеличению затрат на обслуживание в ландшафтных условиях, если кто-то хочет, чтобы пальма выглядела «чисто», которая не самоочищается.
Кокосовые пальмы были очень полезны гавайцам (пища, волокно, ткань, солома, древесина) и встречаются во всех тропиках. Люди, посещающие тропики, ожидают их увидеть (, рис. 36 и 37, ).
К сожалению, кокосовые пальмы могут быть опасны для неосторожных людей под ними. Тяжёлый плод может упасть в любой момент и ранить или даже убить тех, кому посчастливилось попасть под удар.Во время урагана кокосы могут превратиться в смертоносные летящие по ветру ракеты. В городах, где кокосовые пальмы являются частью городского пейзажа, кокосы регулярно обрезают с деревьев, чтобы предотвратить возможные травмы от падающих кокосов, а под кокосовыми пальмами в общественных местах часто размещают предупреждающие знаки. Кокосы распространяются по тропикам океанскими течениями (и людьми — их завезли на Гавайи полинезийцы = растения-каноэ). Кокос не прорастет, пока он плавает, потому что «вода» внутри него не должна двигаться, чтобы он пророс.Таким образом, он должен быть выброшен на берег и некоторое время лежать неподвижно, прежде чем он сможет прорасти. Это важный механизм выживания кокоса. Детеныш/эмбрион кокосового растения находится прямо под одним из 3 «глазков» в «орехе» — глазком, который мягкий. Поскольку этот один глаз мягкий, зародышевый отросток может протиснуться через глазную оболочку и вырасти из твердой скорлупы ореха (, рис. 38 и 39, ).
Кокосы распространяются по тропикам океанскими течениями (и людьми — их завезли на Гавайи полинезийцы = растения-каноэ). Кокос не прорастет, пока он плавает, потому что «вода» внутри него не должна двигаться, чтобы он пророс.Таким образом, он должен быть выброшен на берег и некоторое время лежать неподвижно, прежде чем он сможет прорасти. Это важный механизм выживания кокоса. Детеныш/эмбрион кокосового растения находится прямо под одним из 3 «глазков» в «орехе» — глазком, который мягкий. Поскольку этот один глаз мягкий, зародышевый отросток может протиснуться через глазную оболочку и вырасти из твердой скорлупы ореха (, рис. 38 и 39, ).
К счастью для кокосовой пальмы, она производит как мужские, так и женские цветки в своем соцветии (цветковая структура) и может производить жизнеспособные кокосы (плоды, каждый из которых содержит семя).
Приходите во Всемирный ботанический сад и посмотрите на все это и многое другое во время одной из наших веселых и захватывающих экскурсий по саду!
Доктор Лэнни Нил, почетный директор сада, Всемирный ботанический сад
анатомия пальм (Arecaceae–Palmae) | Анналы ботаники
Пальмы, без сомнения, являются одним из самых важных семейств растений с точки зрения человека, и они жизненно важны как для экологического, так и для экономического благополучия тропических регионов мира. Поэтому понимание того, как они функционируют, является важной задачей для биологов пальм, которые будут рады появлению этого нового и важного учебника Томлинсона, Хорна и Фишера.
Поэтому понимание того, как они функционируют, является важной задачей для биологов пальм, которые будут рады появлению этого нового и важного учебника Томлинсона, Хорна и Фишера.
Анатомия ладоней делает то, что написано на обложке: читателю предоставляется подробный обзор анатомического разнообразия Arecaceae. Эта информация получает филогенетический акцент благодаря включению в качестве фона к анатомическим данным хорошо установленной в настоящее время филогении семейства на уровне рода.
Пальмы — легко узнаваемая группа растений. Здесь особенно важны две их типичные характеристики: во-первых, их большой размер создает проблемы для микроскопических исследований и требует, как отмечают авторы, «кажущегося редукционистским» подхода с привлечением «небольших образцов из относительно стандартных областей растения». Во-вторых, ткани ладони часто очень волокнистые, с сильным одревеснением и частым появлением кремнеземных тел, что затрудняет их фиксацию и разрез.Как и следовало ожидать от группы признанных лидеров в области анатомии ладони, авторы с большим успехом преодолели эти препятствия. Многие изображения в книге были получены из невстроенного материала и подготовлены специально для этой публикации.
Многие изображения в книге были получены из невстроенного материала и подготовлены специально для этой публикации.
Книга состоит из 276 страниц и представляет собой произведение поразительной красоты, содержащее около 800 фотографий, подавляющее большинство из которых цветные. Он состоит из различных разделов: короткая глава об используемых анатомических методах; краткий обзор биологии пальмы; главы о строении листовой пластинки, оси листа, стебля, корня и сосудистых тканей; затем обзор анатомической эволюции ладоней.Оставшаяся (и большая) часть книги посвящена обсуждению анатомических особенностей семейства пальмовых в филогенетическом контексте. Рассматривается каждый из признанных в настоящее время 183 родов пальм. Рассмотрено внутреннее строение всех вегетативных органов ладоней с особым акцентом на анатомию пластинки. Таксономические ключи, основанные на анатомических признаках, предоставляются для групп на разных уровнях семьи. Таким образом, ключи даны для некоторых родов (например, Hyophorbe ), в то время как другие относятся к подтрибам (например,г. Attaleinae) или трибы (например, Ceroxyleae).
Attaleinae) или трибы (например, Ceroxyleae).
Хотя авторы подчеркивают в предисловии, что следует признать «незавершенность» этой работы, большинству читателей сразу станет ясно, что это титанический труд, который еще долгие годы будет служить ориентиром для пальмового сообщества . Анатомия пальм станет обязательным справочником не только для биологов пальм, но и для более широкого сообщества структурных биологов растений, интересующихся разнообразием и эволюцией анатомических признаков.Он также станет идеальным дополнением к «Структурная биология пальм» , еще одной существенной, но более общей работе, опубликованной ранее П. Барри Томлинсоном (1990), последняя теперь доступна в версии «печать по требованию».
ЦИТИРОВАННАЯ ЛИТЕРАТУРА
. ,Структурная биология пальм
,1990
Oxford
Oxford University Press
© The Author, 2011. Опубликовано издательством Oxford University Press от имени компании Annals of Botany. Все права защищены. Для получения разрешений отправьте электронное письмо по адресу: [email protected]
Все права защищены. Для получения разрешений отправьте электронное письмо по адресу: [email protected]
Структура и развитие цветка однодомной пальмы Gaussia attenuata (Arecaceae; Arecoideae) | Анналы ботаники
Аннотация
Исходная информация и цели
Половой диморфизм как на уровне цветков, так и на уровне растений широко распространен в семействе пальмовых (Arecaceae), в отличие от покрытосеменных растений в целом. Триба Chamaedoreeae представляет особый интерес для изучения эволюции сексуального самовыражения, так как двудомность, по-видимому, дважды развилась в этой группе независимо от однодомного предка.Чтобы понять основные эволюционные пути, важно получить подробную информацию о структуре и развитии цветка в каждой из основных клад.
Методы
Препарирование и световая и сканирующая электронная микроскопия были выполнены на развивающихся цветках Gaussia attenuata , неотропического вида, принадлежащего к одному из трех однодомных родов трибы.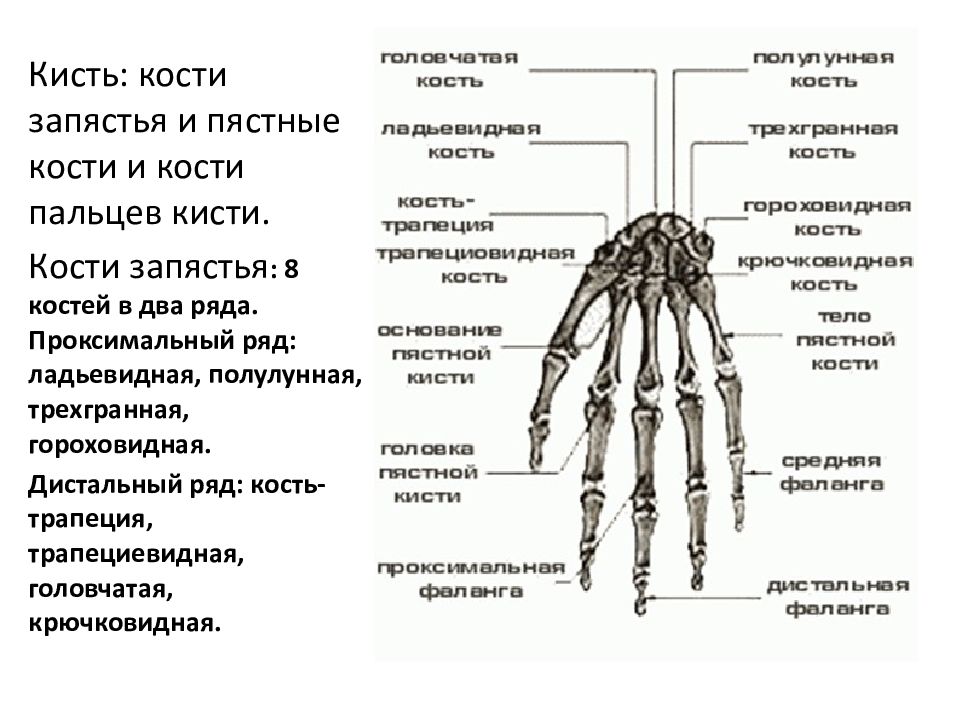
Основные результаты
Аналогичные виды других однодомных родов Chamaedoreeae (а именно Hyophorbe и Synechanthus ), G.attenuata образует группу обоеполых цветков, известную как ацервулюс, состоящую из ряда мужских цветков с базальным женским цветком. В то время как стерильный андроцей женских цветков заканчивал свое развитие на ранней стадии онтогенеза цветка, пестик мужских цветков был крупного размера, но без узнаваемой семяпочки, развиваясь в течение более длительного периода времени. Заметная нектарная дифференциация в пестичных ножках предполагает возможную роль в привлечении опылителей.
Выводы
Gaussia attenuata демонстрирует ряд цветочных признаков, которые, вероятно, являются предками трибы, в частности соцветие acervulus, которое сохраняется у других однодомных родов, а также (хотя и в однополой мужской форме) у двудомные роды ( Wendlandiella и несколько видов Chamaedorea ).Сравнение с более ранними данными по другим родам позволяет предположить, что крупные нектароносные пестики и ранняя остановка развития стаминодий также могут рассматриваться как предковые признаки этой трибы.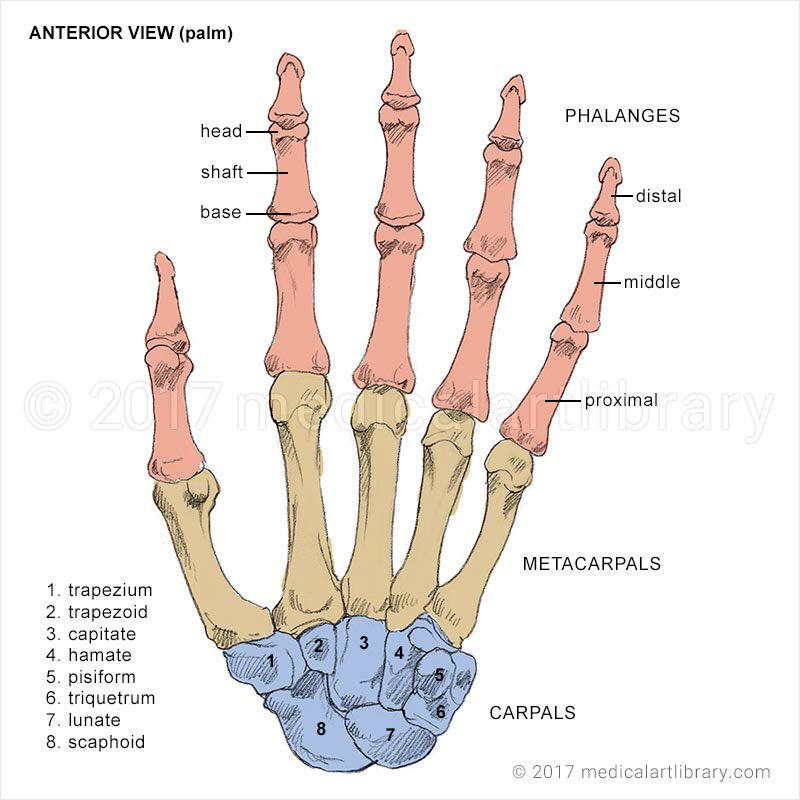
ВВЕДЕНИЕ
Структура цветка является ключевым морфологическим признаком, определяющим успешность цветения растений. Многие цветочные признаки демонстрируют сложные эволюционные модели, отражающие большое разнообразие покрытосеменных растений, ярким примером которых является сексуальное выражение и его разнообразные формы.Хотя считается вероятным, что предковый цветок покрытосеменных растений был обоеполым, остаются некоторые сомнения (Endress and Doyle, 2009). Считается, что гермафродитизм (производство одного типа цветков с функциональными мужскими и женскими репродуктивными органами) составляет 85–90 % всех покрытосеменных растений (Endress and Doyle, 2009; Diggle et al. , 2011). Оставшееся меньшинство видов демонстрирует либо однодомность (отдельные мужские и женские цветки на одном и том же растении), двудомность (отдельные половые растения), либо промежуточные формы, часто называемые полигамией.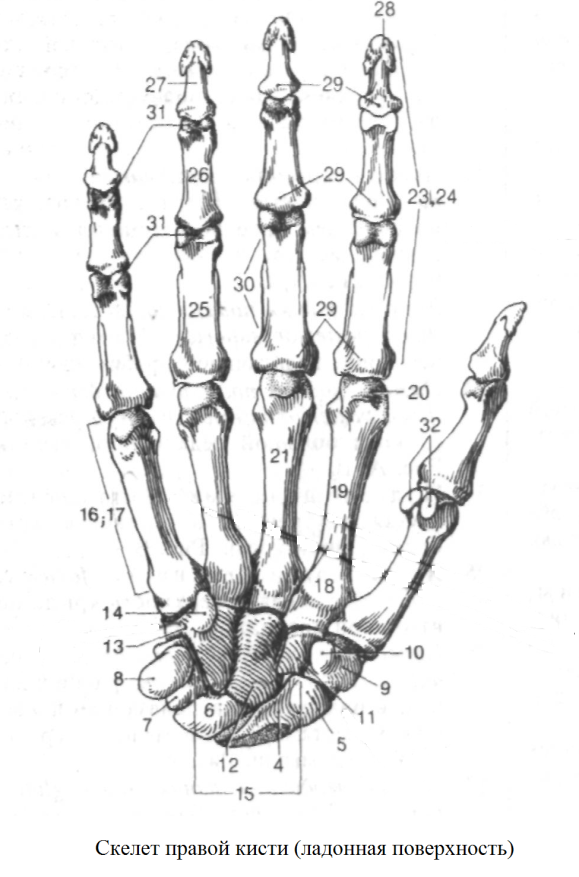 Считается, что половой диморфизм обеспечивает средства для оптимизации распределения ресурсов для воспроизводства в растении (Sánchez and Pannell, 2011) и способствует аутбридингу и гетерозису (Freeman et al. , 1997). Существование отдельных мужских и женских цветков может быть ключевым элементом взаимодействия растений и опылителей (Barrett, 2013).
Считается, что половой диморфизм обеспечивает средства для оптимизации распределения ресурсов для воспроизводства в растении (Sánchez and Pannell, 2011) и способствует аутбридингу и гетерозису (Freeman et al. , 1997). Существование отдельных мужских и женских цветков может быть ключевым элементом взаимодействия растений и опылителей (Barrett, 2013).
Учитывая большое разнообразие форм полового диморфизма, встречающихся у покрытосеменных растений, и сложные филогенетические закономерности, лежащие в основе их распространения, существует значительный интерес в сосредоточении внимания на конкретных кладах, которые могут служить в качестве тематических исследований эволюции.Особенно актуальной группой в этом контексте является семейство пальмовых (Arecaceae). Это семейство включает 185 родов и около 2500 видов (Govaerts, Dransfield, 2005; Dransfield et al. , 2008), обитающих в основном в тропиках. Цветочный половой диморфизм особенно распространен у пальм, для которых он является скорее правилом, чем исключением; в отличие от покрытосеменных в целом, примерно четыре пятых родов пальм двудомные, однодомные или полигамные (Dransfield et al. , 2008).Наиболее распространенным типом половой системы является моноэция, которая встречается примерно у 65 % родов пальм (Dransfield et al. , 2008) и доминирует в самом крупном подсемействе Arecoideae, внутри которого только одна из 14 триб неоднородна. однодомный. Исключением является триба Chamaedoreeae, образующая монофилетическую группу из пяти родов, два из которых двудомные ( Chamaedorea и Wendlandiella ), а остальные однодомные ( Gaussia , Hyophorbe и Synechanthus ).Триба включает наиболее богатый видами род пальм неотропиков, Chamaedorea , который демонстрирует высокую изменчивость вегетативной и репродуктивной морфологии. Более того, это племя демонстрирует одно из самых необычных расхождений в семействе пальмовых: Hyophorbe эндемичны для Маскаренских островов, а другие роды распространены в северной части Южной Америки, Центральной Америке и Карибском бассейне (Cuenca et al. , 2008; Dransfield и др.
, 2008).Наиболее распространенным типом половой системы является моноэция, которая встречается примерно у 65 % родов пальм (Dransfield et al. , 2008) и доминирует в самом крупном подсемействе Arecoideae, внутри которого только одна из 14 триб неоднородна. однодомный. Исключением является триба Chamaedoreeae, образующая монофилетическую группу из пяти родов, два из которых двудомные ( Chamaedorea и Wendlandiella ), а остальные однодомные ( Gaussia , Hyophorbe и Synechanthus ).Триба включает наиболее богатый видами род пальм неотропиков, Chamaedorea , который демонстрирует высокую изменчивость вегетативной и репродуктивной морфологии. Более того, это племя демонстрирует одно из самых необычных расхождений в семействе пальмовых: Hyophorbe эндемичны для Маскаренских островов, а другие роды распространены в северной части Южной Америки, Центральной Америке и Карибском бассейне (Cuenca et al. , 2008; Dransfield и др.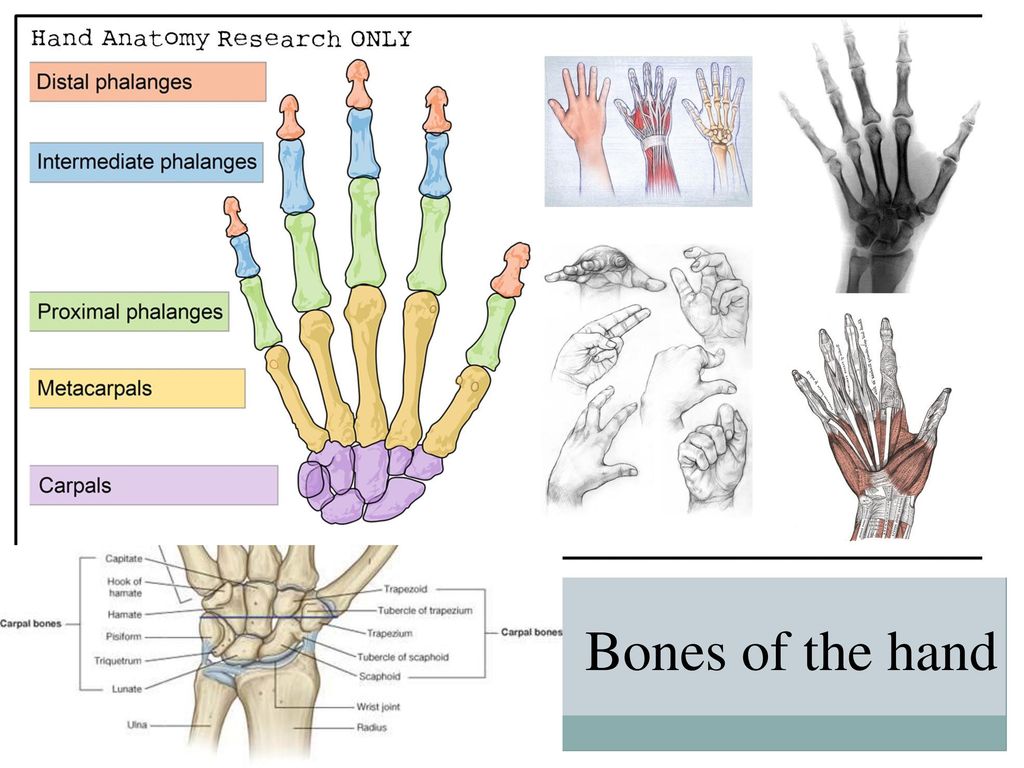 , 2008).Триба Chamaedoreeae была определена как монофилетическая с высокой поддержкой в нескольких филогенетических исследованиях, основанных на морфологических и молекулярных данных (Uhl et al. , 1995; Hahn, 2002; Asmussen et al. , 2006; Cuenca and Asmussen, 2007; Cuenca и др. , 2008, 2009; Baker и др. , 2009, 2011). Возможные филогенетические отношения родов компонентов, полученные в результате вышеупомянутых исследований, схематически представлены на рис. 1А. Включение Chamaedoreeae в подсемейство Arecoideae (Dransfield et al., 2005; Асмуссен и др. , 2006), в отличие от более ранней классификации Ceroxyloideae (например, Uhl and Dransfield, 1987; Dransfield and Uhl, 1998), поднимает эволюционные вопросы, касающиеся полового выражения и расположения цветков внутри соцветия. У подавляющего большинства родов пальм цветки сгруппированы в симподиальные грозди или цинцинны. Большинство видов подсемейства Arecoideae образуют цветки, сгруппированные в триады или группы, уменьшенные по сравнению с этой предполагаемой предковой формой (Moore, 1973; Uhl and Dransfield, 1987; Dransfield and Uhl, 1998; Dransfield et al.
, 2008).Триба Chamaedoreeae была определена как монофилетическая с высокой поддержкой в нескольких филогенетических исследованиях, основанных на морфологических и молекулярных данных (Uhl et al. , 1995; Hahn, 2002; Asmussen et al. , 2006; Cuenca and Asmussen, 2007; Cuenca и др. , 2008, 2009; Baker и др. , 2009, 2011). Возможные филогенетические отношения родов компонентов, полученные в результате вышеупомянутых исследований, схематически представлены на рис. 1А. Включение Chamaedoreeae в подсемейство Arecoideae (Dransfield et al., 2005; Асмуссен и др. , 2006), в отличие от более ранней классификации Ceroxyloideae (например, Uhl and Dransfield, 1987; Dransfield and Uhl, 1998), поднимает эволюционные вопросы, касающиеся полового выражения и расположения цветков внутри соцветия. У подавляющего большинства родов пальм цветки сгруппированы в симподиальные грозди или цинцинны. Большинство видов подсемейства Arecoideae образуют цветки, сгруппированные в триады или группы, уменьшенные по сравнению с этой предполагаемой предковой формой (Moore, 1973; Uhl and Dransfield, 1987; Dransfield and Uhl, 1998; Dransfield et al.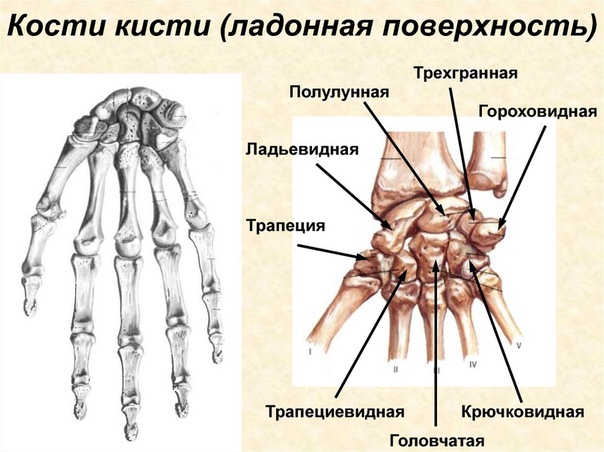 , 2008). Триада представляет собой симподиальное соцветие, состоящее из двух боковых тычиночных цветков и центрального пестичного цветка с тремя ассоциированными прицветниками (Uhl, 1976; Ortega and Stauffer, 2011). Chamaedoreeae — единственная триба в Arecoideae, которая отклоняется от этой общей модели, производя цветы, расположенные в характерной структуре, известной как acervulus (Uhl and Moore, 1978; Uhl and Dransfield, 1987; Ortega and Stauffer, 2011). Acervulus состоит, в случае однодомных родов, из переменного числа мужских цветков, расположенных в ряд, который заканчивается базальным женским цветком.В раздельнополых родах ( Chamaedorea и Wendlandiella ) ацервулии, состоящие только из мужских цветков, наблюдаются у некоторых видов, другие дают одиночные мужские цветки. Хотя особенности половой системы и соцветий считались важными при выводе об эволюционных отношениях между родами Chamaedoreeae, недавние исследования показали, что внутри трибы происходили множественные переходы между предполагаемыми наследственными признаками однодомности и образования ацервулуса, с одной стороны, и производными состояниями.
, 2008). Триада представляет собой симподиальное соцветие, состоящее из двух боковых тычиночных цветков и центрального пестичного цветка с тремя ассоциированными прицветниками (Uhl, 1976; Ortega and Stauffer, 2011). Chamaedoreeae — единственная триба в Arecoideae, которая отклоняется от этой общей модели, производя цветы, расположенные в характерной структуре, известной как acervulus (Uhl and Moore, 1978; Uhl and Dransfield, 1987; Ortega and Stauffer, 2011). Acervulus состоит, в случае однодомных родов, из переменного числа мужских цветков, расположенных в ряд, который заканчивается базальным женским цветком.В раздельнополых родах ( Chamaedorea и Wendlandiella ) ацервулии, состоящие только из мужских цветков, наблюдаются у некоторых видов, другие дают одиночные мужские цветки. Хотя особенности половой системы и соцветий считались важными при выводе об эволюционных отношениях между родами Chamaedoreeae, недавние исследования показали, что внутри трибы происходили множественные переходы между предполагаемыми наследственными признаками однодомности и образования ацервулуса, с одной стороны, и производными состояниями.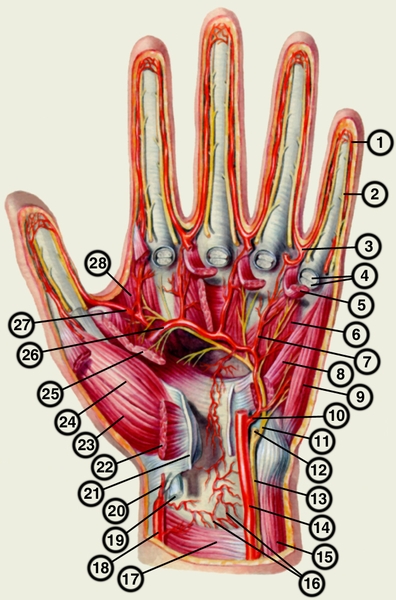 раздельнополых и одиночных цветков с другой (Thomas et al., 2006; Куэнка и Асмуссен, 2007 г.; Куэнка и др. , 2009 г.).
раздельнополых и одиночных цветков с другой (Thomas et al., 2006; Куэнка и Асмуссен, 2007 г.; Куэнка и др. , 2009 г.).
Рис. 1.
Схематическая филогения трибы Chamaedoreeae и взрослой особи G. attenuata . (A) Возможные филогенетические отношения между родами трибы Chamaedoreeae, основанные на исследованиях Cuenca et al . (2008). (B) Взрослый экземпляр Gaussia attenuata (фото любезно предоставлено Л. Нобликом, Ботанический центр Монтгомери).
Рис.1.
Схематическая филогения трибы Chamaedoreeae и взрослой особи G. attenuata . (A) Возможные филогенетические отношения между родами трибы Chamaedoreeae, основанные на исследованиях Cuenca et al . (2008). (B) Взрослый экземпляр Gaussia attenuata (фото любезно предоставлено Л. Нобликом, Ботанический центр Монтгомери).
Исследования развития ряда видов пальм привели к пониманию онтогенеза и разнообразия соцветий (Uhl and Dransfield, 1984). Упомянутая выше структура цинциннуса представляет собой монохазиум, в котором цветки следуют зигзагообразно вдоль одной стороны оси (Rickett, 1955). Этот тип роста дает цветочные кисти различной формы. Acervulus, производимый членами трибы Chamaedoreeae, в настоящее время считается перевернутым цинциннусом, состоящим из отдельных линий тесно расположенных двурядных однополых сидячих цветков, сросшихся с рахиллами (Uhl and Moore, 1978; Uhl and Dransfield, 1987; Dransfield ). и др. , 2008; Ortega and Stauffer, 2011).Проксимальный цветок обычно женский, а дистальные — мужские, с редуцированными прицветниками или незаметными при цветении. Acervuli демонстрируют почти исключительно протандрический характер цветочного созревания. У некоторых однодомных видов сообщалось о случайном появлении на концах рахилл ацервули, состоящих только из мужских цветков, или даже об образовании одиночных мужских цветков (Quero, 1982; Quero and Read, 1986; Ortega and Stauffer, 2011).
Упомянутая выше структура цинциннуса представляет собой монохазиум, в котором цветки следуют зигзагообразно вдоль одной стороны оси (Rickett, 1955). Этот тип роста дает цветочные кисти различной формы. Acervulus, производимый членами трибы Chamaedoreeae, в настоящее время считается перевернутым цинциннусом, состоящим из отдельных линий тесно расположенных двурядных однополых сидячих цветков, сросшихся с рахиллами (Uhl and Moore, 1978; Uhl and Dransfield, 1987; Dransfield ). и др. , 2008; Ortega and Stauffer, 2011).Проксимальный цветок обычно женский, а дистальные — мужские, с редуцированными прицветниками или незаметными при цветении. Acervuli демонстрируют почти исключительно протандрический характер цветочного созревания. У некоторых однодомных видов сообщалось о случайном появлении на концах рахилл ацервули, состоящих только из мужских цветков, или даже об образовании одиночных мужских цветков (Quero, 1982; Quero and Read, 1986; Ortega and Stauffer, 2011).
Что касается эволюции сексуального самовыражения, то однодомность и двудомность, по-видимому, неоднократно развивались у пальм независимо друг от друга (Weiblen et al.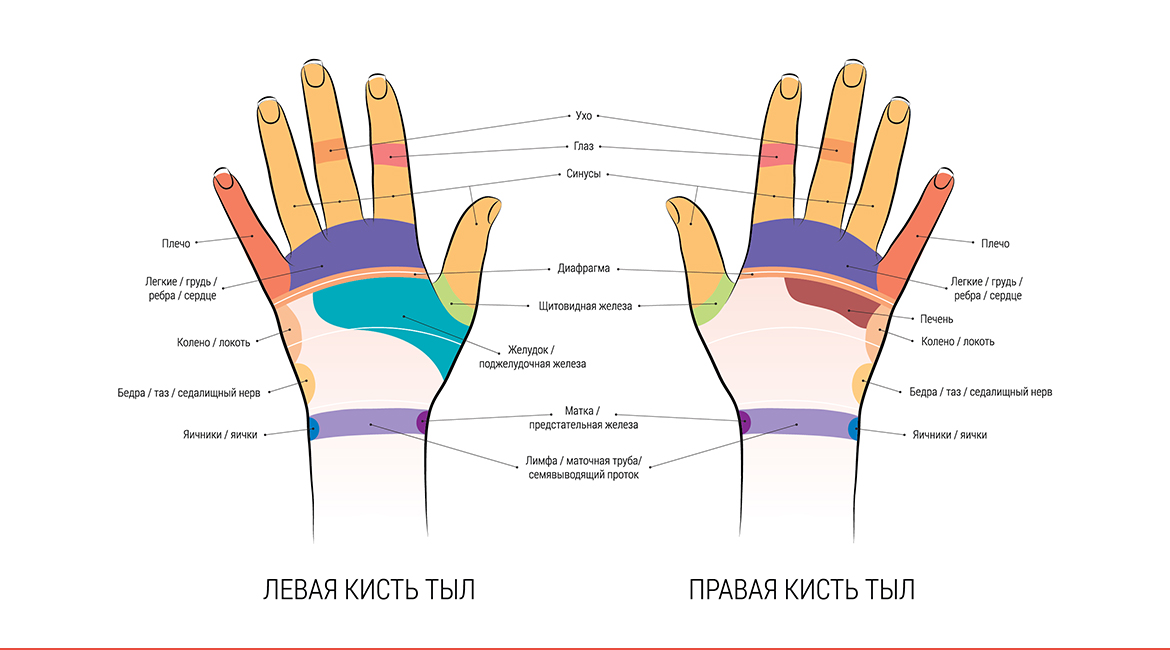 , 2000). Подсчитано, что двудомность появлялась по крайней мере девять раз в ходе их эволюции от однодомных или гермафродитных предков. Очень мало известно о механизмах, с помощью которых пальмы и их цветы приобретают мужскую или женскую идентичность, однако существует большой интерес к изучению этого явления, поскольку это улучшит наше понимание эволюции пальмового цветка и предоставит знания для повышения продуктивности растений. культивируемых пальм и получить новое представление о динамике популяций видов, находящихся под угрозой исчезновения.В этом контексте замечательные особенности трибы Chamaedoreeae в отношении паттернов полового выражения и других цветочных признаков побудили нас исследовать черты развития и строения одного из составляющих его родов, чтобы улучшить наши знания о сексуальности в семействе пальмовых. целое. Если мы хотим понять, как развивались цветочные признаки внутри и за пределами Chamaedoreeae, сначала необходимо получить подробные сведения о цветочной структуре и развитии во всех основных кладах.
, 2000). Подсчитано, что двудомность появлялась по крайней мере девять раз в ходе их эволюции от однодомных или гермафродитных предков. Очень мало известно о механизмах, с помощью которых пальмы и их цветы приобретают мужскую или женскую идентичность, однако существует большой интерес к изучению этого явления, поскольку это улучшит наше понимание эволюции пальмового цветка и предоставит знания для повышения продуктивности растений. культивируемых пальм и получить новое представление о динамике популяций видов, находящихся под угрозой исчезновения.В этом контексте замечательные особенности трибы Chamaedoreeae в отношении паттернов полового выражения и других цветочных признаков побудили нас исследовать черты развития и строения одного из составляющих его родов, чтобы улучшить наши знания о сексуальности в семействе пальмовых. целое. Если мы хотим понять, как развивались цветочные признаки внутри и за пределами Chamaedoreeae, сначала необходимо получить подробные сведения о цветочной структуре и развитии во всех основных кладах.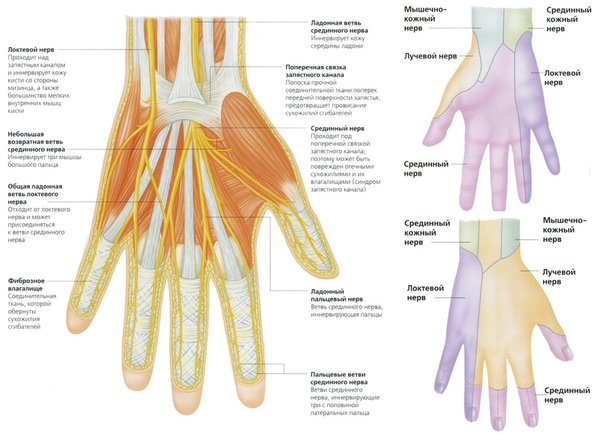 Более ранние исследования предоставили данные о морфологии и анатомии цветков родов Hyophorbe и Chamaedorea (Uhl and Moore, 1971, 1978; Uhl, 1978; Askgaard et al. , 2008; Ortega and Stauffer, 2011), с небольшим информация пока доступна для Synechanthus , Wendlandiella и Gaussia . Здесь мы описываем исследования развития цветков и ацервулюса у вида Gaussia attenuata , однодомной одиночной пальмы среднего размера (рис.1B), эндемичен для Пуэрто-Рико (Quero and Read, 1986; Henderson et al. , 1995) и известен под общим названием пальма Llume. Gaussia оказался монофилетическим с высокой поддержкой (Cuenca and Asmussen, 2007), род, состоящий из пяти признанных видов (Quero and Read, 1986; Moya et al. , 1991). Основываясь на сходстве вегетативного состояния с Pseudophoenix (представителем сестринского подсемейства Ceroxyloideae), Уль и Дрансфилд (1987) сочли Gaussia наименее специализированным родом трибы и указали на общие признаки с Hyophorbe и .
Более ранние исследования предоставили данные о морфологии и анатомии цветков родов Hyophorbe и Chamaedorea (Uhl and Moore, 1971, 1978; Uhl, 1978; Askgaard et al. , 2008; Ortega and Stauffer, 2011), с небольшим информация пока доступна для Synechanthus , Wendlandiella и Gaussia . Здесь мы описываем исследования развития цветков и ацервулюса у вида Gaussia attenuata , однодомной одиночной пальмы среднего размера (рис.1B), эндемичен для Пуэрто-Рико (Quero and Read, 1986; Henderson et al. , 1995) и известен под общим названием пальма Llume. Gaussia оказался монофилетическим с высокой поддержкой (Cuenca and Asmussen, 2007), род, состоящий из пяти признанных видов (Quero and Read, 1986; Moya et al. , 1991). Основываясь на сходстве вегетативного состояния с Pseudophoenix (представителем сестринского подсемейства Ceroxyloideae), Уль и Дрансфилд (1987) сочли Gaussia наименее специализированным родом трибы и указали на общие признаки с Hyophorbe и . Синехант .Самые последние филогенетические анализы не дали однозначного прояснения взаимоотношений между пятью родами Chamaedoreeae (рис. 1А; Thomas et al. , 2006; Cuenca and Asmussen, 2007; Cuenca et al. , 2009; Baker и др. , 2011). Тем не менее, Gaussia была обнаружена как сестра Chamaedorea с умеренной или высокой степенью поддержки в анализах, основанных на морфологических или молекулярных данных (Thomas et al. , 2006; Cuenca and Asmussen, 2007; Cuenca et al., 2008, 2009; Бейкер и др. , 2011). Независимо от точной филогенетической топологии трибы, род Gaussia составляет важную часть мозаики, которую необходимо собрать, чтобы понять, как эволюционировали цветочные признаки у Chamaedoreeae и родственных клад. Представленные здесь данные были получены с целью заполнения этого заметного пробела в современных знаниях.
Синехант .Самые последние филогенетические анализы не дали однозначного прояснения взаимоотношений между пятью родами Chamaedoreeae (рис. 1А; Thomas et al. , 2006; Cuenca and Asmussen, 2007; Cuenca et al. , 2009; Baker и др. , 2011). Тем не менее, Gaussia была обнаружена как сестра Chamaedorea с умеренной или высокой степенью поддержки в анализах, основанных на морфологических или молекулярных данных (Thomas et al. , 2006; Cuenca and Asmussen, 2007; Cuenca et al., 2008, 2009; Бейкер и др. , 2011). Независимо от точной филогенетической топологии трибы, род Gaussia составляет важную часть мозаики, которую необходимо собрать, чтобы понять, как эволюционировали цветочные признаки у Chamaedoreeae и родственных клад. Представленные здесь данные были получены с целью заполнения этого заметного пробела в современных знаниях.
МАТЕРИАЛЫ И МЕТОДЫ
Растительный материал
Морфо-анатомические и онтогенетические исследования структуры цветка Gaussia attenuata проводились на цветках на разных стадиях развития от ранних цветочных почек до цветков в стадии цветения или близко к нему.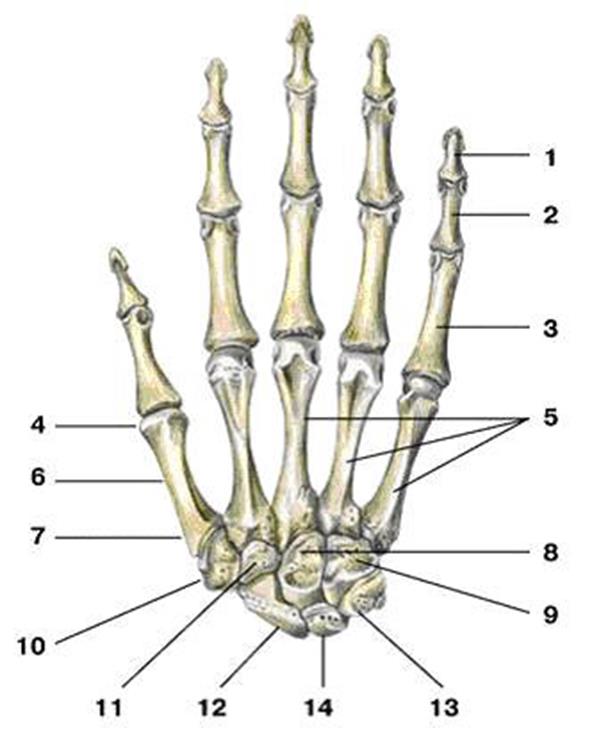 Цветы собраны из живой коллекции Ботанического центра Монтгомери (MBC) в Майами (США). Исследование было ограничено количеством доступных экземпляров, наличием соцветий на подходящей стадии развития и необходимостью получения цветков без летальных последствий для особи, которые могут возникнуть при обширном рассечении пальмы. Фиксацию проводили, как описано ранее (Adam et al. ., 2005). У некоторых цветков были удалены чашелистики и лепестки, чтобы облегчить наблюдение за внутренними органами.
Цветы собраны из живой коллекции Ботанического центра Монтгомери (MBC) в Майами (США). Исследование было ограничено количеством доступных экземпляров, наличием соцветий на подходящей стадии развития и необходимостью получения цветков без летальных последствий для особи, которые могут возникнуть при обширном рассечении пальмы. Фиксацию проводили, как описано ранее (Adam et al. ., 2005). У некоторых цветков были удалены чашелистики и лепестки, чтобы облегчить наблюдение за внутренними органами.
Морфологический анализ
цветка препарировали для морфологического исследования под стереомикроскопом (Wild M3B). Некоторые части препарированных образцов были отобраны для сканирующей электронной микроскопии (СЭМ). Они были обезвожены, высушены в критической точке и покрыты золотым напылением в Музее естественной истории де ла Виль де Женев (Женева, Швейцария) и в Центре ресурсов и клеточных изображений (CRIC, Монпелье, Франция). Микрофотографии были получены с использованием сканирующего электронного микроскопа JEOL (JSM-6510LV) в Центре биовизуализации факультета наук (Женевский университет) и сканирующего электронного микроскопа Hitachi S4000 в лаборатории CRIC.
Микрофотографии были получены с использованием сканирующего электронного микроскопа JEOL (JSM-6510LV) в Центре биовизуализации факультета наук (Женевский университет) и сканирующего электронного микроскопа Hitachi S4000 в лаборатории CRIC.
Анатомический анализ
Детальная анатомо-онтогенетическая характеристика соцветий и цветков G. attenuata проведена методом заливки в парафин с последующим окрашиванием срезов астра синим и фуксином и посевом на среду Юкитта в лаборатории гистотехнологии биологического факультета Национальный университет Колумбии. Микрофотографии анатомических срезов были получены с использованием микроскопа Leica DMIRE2 и камеры Leica DC 300F.
РЕЗУЛЬТАТЫ
Развитие ацервулюса
Соцветие G. attenuata было межлистным (рис. 1Б). Его общая длина составляла ∼1 м, около половины которой приходилось на стебель. Характеризуется наличием от четырех до семи стойких трубчатых черешковых прицветников. В основном он был разветвлен на два порядка, хотя было замечено, что только ветви первого порядка образуются в средней и дистальной частях. Каждая рахилла несла многочисленные спирально расположенные ацервулы (рис.2А показана группа незрелых ацервулиев, обычно состоящая из четырех однополых цветков, хотя в некоторых случаях наблюдалось пять цветков. Только один из этих цветков был женским (см. далее). Каждая ацервулюс была окружена очень коротким прицветником, и именно в пазухе этого прицветника зародились все цветки (рис. 2B). Мужские цветки были инициированы последовательно в базипетальном образце, который завершился инициацией женского цветка (рис. 2C). В каждой ацервулюсе цветки располагались зигзагообразно, хотя до цветения они имели поверхностный вид линейного расположения (рис.2А, С).
Его общая длина составляла ∼1 м, около половины которой приходилось на стебель. Характеризуется наличием от четырех до семи стойких трубчатых черешковых прицветников. В основном он был разветвлен на два порядка, хотя было замечено, что только ветви первого порядка образуются в средней и дистальной частях. Каждая рахилла несла многочисленные спирально расположенные ацервулы (рис.2А показана группа незрелых ацервулиев, обычно состоящая из четырех однополых цветков, хотя в некоторых случаях наблюдалось пять цветков. Только один из этих цветков был женским (см. далее). Каждая ацервулюс была окружена очень коротким прицветником, и именно в пазухе этого прицветника зародились все цветки (рис. 2B). Мужские цветки были инициированы последовательно в базипетальном образце, который завершился инициацией женского цветка (рис. 2C). В каждой ацервулюсе цветки располагались зигзагообразно, хотя до цветения они имели поверхностный вид линейного расположения (рис.2А, С).
Рис. 2.
СЭМ и макроскопические изображения развивающейся ацервулюса у Gaussia attenuata . (A) Часть рахиллы, показывающая ацервули со всеми цветками в бутонах (этап 1; таблица 1). (B) Женская цветочная почка и стягивающий прицветник. (C) Acervulus со всеми цветками в бутонах (этап 1; таблица 1). (D) Часть рахиллы, показывающая ацервули с одним мужским цветком при цветении и двумя мужскими цветками плюс женский цветок в бутоне (этап 2; таблица 1). (E, F) Часть рахиллы, показывающая ацервулюс с двумя мужскими цветками и женским цветком в бутоне (этап 3; таблица 1).Обратите внимание, что лепестки (E) были удалены, а один мужской цветок уже осыпался. (G, H) Часть рахиллы, показывающая ацервулюс с одним мужским цветком при цветении и одним мужским цветком плюс женский цветок в бутоне (этап 4; таблица 1). (I, J) Часть рахиллы, показывающая ацервулюс с одним мужским цветком в стадии цветения и женским цветком в бутоне (этап 5; таблица 1). Обратите внимание, что лепестки были удалены в I. (K, L) Часть рахиллы, показывающая женский цветок при цветении. Шкала баров: (А) = 5 мм; (В) = 200 мкм; (С) = 1 мм; (Д) = 5 мм; (Е) = 1 мм; (F) = 2 мм; (Г) = 1 мм; (Н) = 5 мм; (I) = 1 мм; (Дж) = 1 мм; (К) = 1 мм; (L) = 500 мкм.
(A) Часть рахиллы, показывающая ацервули со всеми цветками в бутонах (этап 1; таблица 1). (B) Женская цветочная почка и стягивающий прицветник. (C) Acervulus со всеми цветками в бутонах (этап 1; таблица 1). (D) Часть рахиллы, показывающая ацервули с одним мужским цветком при цветении и двумя мужскими цветками плюс женский цветок в бутоне (этап 2; таблица 1). (E, F) Часть рахиллы, показывающая ацервулюс с двумя мужскими цветками и женским цветком в бутоне (этап 3; таблица 1).Обратите внимание, что лепестки (E) были удалены, а один мужской цветок уже осыпался. (G, H) Часть рахиллы, показывающая ацервулюс с одним мужским цветком при цветении и одним мужским цветком плюс женский цветок в бутоне (этап 4; таблица 1). (I, J) Часть рахиллы, показывающая ацервулюс с одним мужским цветком в стадии цветения и женским цветком в бутоне (этап 5; таблица 1). Обратите внимание, что лепестки были удалены в I. (K, L) Часть рахиллы, показывающая женский цветок при цветении. Шкала баров: (А) = 5 мм; (В) = 200 мкм; (С) = 1 мм; (Д) = 5 мм; (Е) = 1 мм; (F) = 2 мм; (Г) = 1 мм; (Н) = 5 мм; (I) = 1 мм; (Дж) = 1 мм; (К) = 1 мм; (L) = 500 мкм.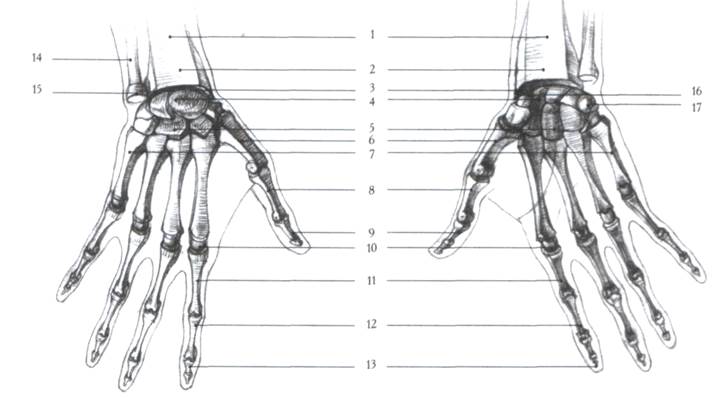 Br, прицветник; Са, плодолистик; Fb, женская цветочная почка; M1, первый мужской цветок при цветении; M2, второй мужской цветок при цветении; M3, третий мужской цветок при цветении; Mb1, первый мужской цветок в стадии бутона; Mb2, второй мужской цветок в стадии бутона; Mb3, третий мужской цветок в стадии бутона; Mb4, четвертый мужской цветок в стадии бутона; Se1, первый чашелистик; Se2, второй чашелистик; Se3, третий чашелистик; Std, стаминод.
Br, прицветник; Са, плодолистик; Fb, женская цветочная почка; M1, первый мужской цветок при цветении; M2, второй мужской цветок при цветении; M3, третий мужской цветок при цветении; Mb1, первый мужской цветок в стадии бутона; Mb2, второй мужской цветок в стадии бутона; Mb3, третий мужской цветок в стадии бутона; Mb4, четвертый мужской цветок в стадии бутона; Se1, первый чашелистик; Se2, второй чашелистик; Se3, третий чашелистик; Std, стаминод.
Рис. 2.
СЭМ и макроскопические изображения развивающейся ацервулюса у Gaussia attenuata . (A) Часть рахиллы, показывающая ацервули со всеми цветками в бутонах (этап 1; таблица 1).(B) Женская цветочная почка и стягивающий прицветник. (C) Acervulus со всеми цветками в бутонах (этап 1; таблица 1). (D) Часть рахиллы, показывающая ацервули с одним мужским цветком при цветении и двумя мужскими цветками плюс женский цветок в бутоне (этап 2; таблица 1). (E, F) Часть рахиллы, показывающая ацервулюс с двумя мужскими цветками и женским цветком в бутоне (этап 3; таблица 1). Обратите внимание, что лепестки (E) были удалены, а один мужской цветок уже осыпался. (G, H) Часть рахиллы, показывающая ацервулюс с одним мужским цветком при цветении и одним мужским цветком плюс женский цветок в бутоне (этап 4; таблица 1).(I, J) Часть рахиллы, показывающая ацервулюс с одним мужским цветком в стадии цветения и женским цветком в бутоне (этап 5; таблица 1). Обратите внимание, что лепестки были удалены в I. (K, L) Часть рахиллы, показывающая женский цветок при цветении. Шкала баров: (А) = 5 мм; (В) = 200 мкм; (С) = 1 мм; (Д) = 5 мм; (Е) = 1 мм; (F) = 2 мм; (Г) = 1 мм; (Н) = 5 мм; (I) = 1 мм; (Дж) = 1 мм; (К) = 1 мм; (L) = 500 мкм. Br, прицветник; Са, плодолистик; Fb, женская цветочная почка; M1, первый мужской цветок при цветении; M2, второй мужской цветок при цветении; M3, третий мужской цветок при цветении; Mb1, первый мужской цветок в стадии бутона; Mb2, второй мужской цветок в стадии бутона; Mb3, третий мужской цветок в стадии бутона; Mb4, четвертый мужской цветок в стадии бутона; Se1, первый чашелистик; Se2, второй чашелистик; Se3, третий чашелистик; Std, стаминод.
Обратите внимание, что лепестки (E) были удалены, а один мужской цветок уже осыпался. (G, H) Часть рахиллы, показывающая ацервулюс с одним мужским цветком при цветении и одним мужским цветком плюс женский цветок в бутоне (этап 4; таблица 1).(I, J) Часть рахиллы, показывающая ацервулюс с одним мужским цветком в стадии цветения и женским цветком в бутоне (этап 5; таблица 1). Обратите внимание, что лепестки были удалены в I. (K, L) Часть рахиллы, показывающая женский цветок при цветении. Шкала баров: (А) = 5 мм; (В) = 200 мкм; (С) = 1 мм; (Д) = 5 мм; (Е) = 1 мм; (F) = 2 мм; (Г) = 1 мм; (Н) = 5 мм; (I) = 1 мм; (Дж) = 1 мм; (К) = 1 мм; (L) = 500 мкм. Br, прицветник; Са, плодолистик; Fb, женская цветочная почка; M1, первый мужской цветок при цветении; M2, второй мужской цветок при цветении; M3, третий мужской цветок при цветении; Mb1, первый мужской цветок в стадии бутона; Mb2, второй мужской цветок в стадии бутона; Mb3, третий мужской цветок в стадии бутона; Mb4, четвертый мужской цветок в стадии бутона; Se1, первый чашелистик; Se2, второй чашелистик; Se3, третий чашелистик; Std, стаминод.
Как только мужские цветки достигли цветения, их последовательно опадали, так что последним цветком, достигшим цветения, был женский цветок, расположенный проксимально по отношению к рахилле. Наблюдалось, что проксимальные ацервулы демонстрируют более продвинутую стадию развития, чем дистальные. Таким образом, в пределах данного соцветия наблюдался ряд стадий развития цветка в зависимости от положения цветка в ацервулюсе и от положения этого ацервулюса в соцветии. В ацервулах G. наблюдалось шесть ключевых стадий развития.attenuata соцветие (табл. 1). Стадия 1 соответствовала самой ранней наблюдаемой стадии развития, когда все цветки ацервулюса были в бутонах (рис. 2А, С). На стадии 2 acervulus самый дистальный мужской цветок достиг цветения, второй мужской цветок находится на стадии поздней бутоны или только начинает раскрываться, сопровождаемый третьим незрелым мужским цветком и одним незрелым женским цветком (рис. 2D). На стадии 3 один мужской цветок уже осыпался, два оставшихся мужских цветка были еще в стадии бутонизации, а женский цветок был еще закрыт (рис. 2Д, Е). Ацервулюс, определенный как стадия 4, содержал один мужской цветок на стадии цветения и другой мужской цветок на поздней стадии бутона с женским цветком в бутонах (рис. 2G, H). На стадии 5 присутствовал один мужской цветок при цветении, сопровождаемый одним женским цветком на поздней стадии бутонизации, остальные мужские цветки осыпались (рис. 2I, J). На стадии 6 все мужские цветки достигли цветения и опали, остался только зрелый женский цветок (рис. 2K, L).
2Д, Е). Ацервулюс, определенный как стадия 4, содержал один мужской цветок на стадии цветения и другой мужской цветок на поздней стадии бутона с женским цветком в бутонах (рис. 2G, H). На стадии 5 присутствовал один мужской цветок при цветении, сопровождаемый одним женским цветком на поздней стадии бутонизации, остальные мужские цветки осыпались (рис. 2I, J). На стадии 6 все мужские цветки достигли цветения и опали, остался только зрелый женский цветок (рис. 2K, L).
Сводка стадий развития ацервулюса у G.аттенуата
| Стадия . | Мужские цветки в бутоне . | Мужские цветки открытые . | Опадание мужских цветков . | Женские цветки в бутоне . | Женские цветки открытые . | |
|---|---|---|---|---|---|---|
| 1 | 3 | — | — | 1 | — | |
| 2 | 2 | 1 | — | 1 | — | |
| 3 | 2 | — | 1 | 1 | — | |
| 4 | 1 | 1 | 1 | 1 | — | |
| 5 | — | 1 | 2 | 1 | — | |
| 6 | ||||||
| 6 | — | — | — | 3 | — | 1 |
Стадия
.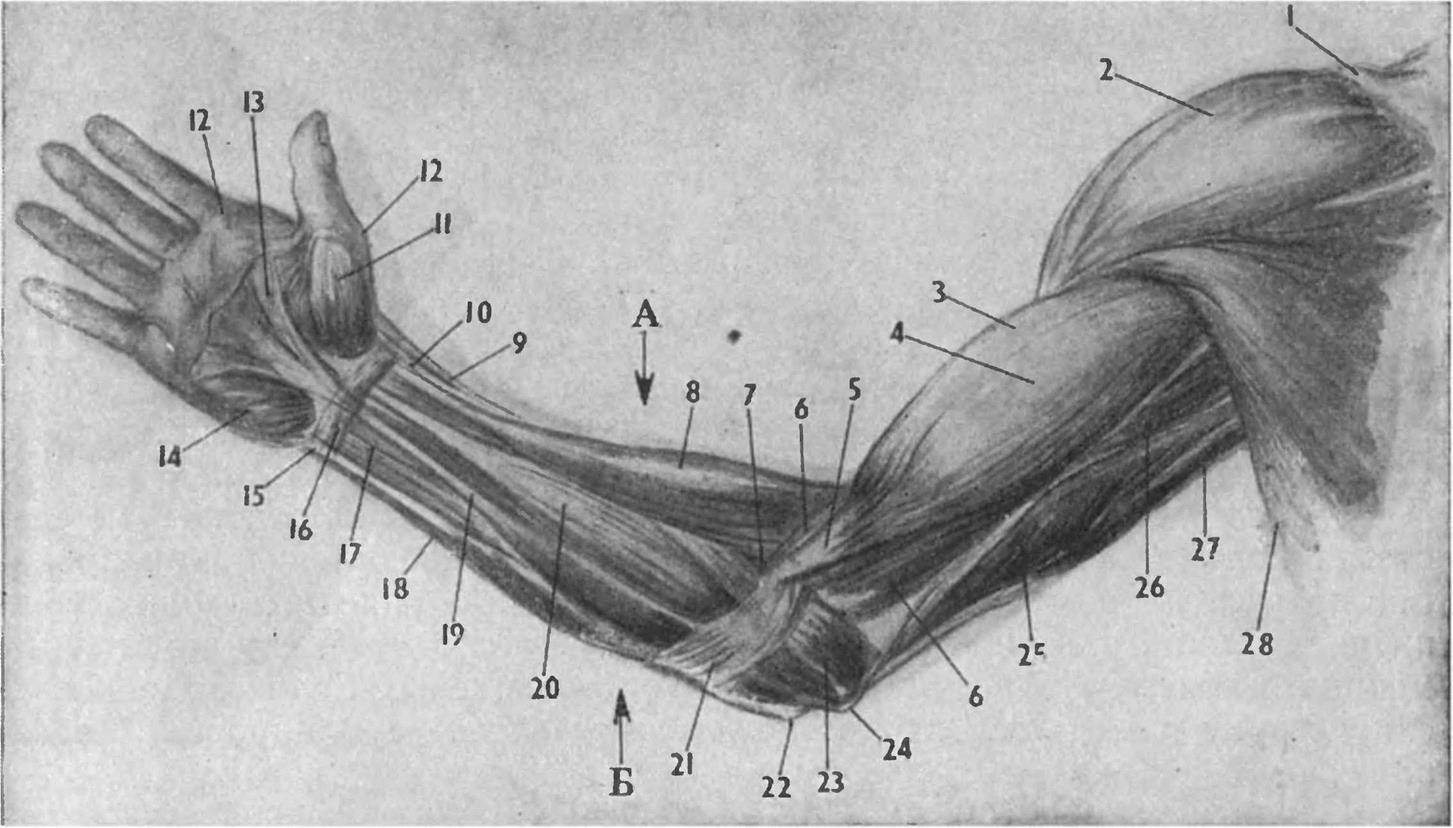 | Мужские цветки в бутоне . | Мужские цветки открытые . | Опадание мужских цветков . | Женские цветки в бутоне . | Женские цветки открытые . | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 3 | — | — | 1 | — | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 2 | 2 | 1 | — | 1 | — | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 3 | 2 | — | 1 | 1 | — | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 4 | 1 | 1 | 1 | 1 | — | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| 5 | — | 1 | 2 | 1 | Таблица
| Стадия . | Мужские цветки в бутоне . | Мужские цветки открытые . | Опадание мужских цветков . | Женские цветки в бутоне
. | Женские цветки открытые . | |
|---|---|---|---|---|---|---|
| 1 | 3 | — | — | 1 | — | |
| 2 | 2 | 1 | — | 1 | — | |
| 3 | 2 | — | 1 | 1 | — | |
| 4 | 1 | 1 | 1 | 1 | — | |
| 5 | — | 1 | 2 | 1 | — | |
| 6 | ||||||
| 6 | — | — | — | 3 | — | 1 |
| Стадия . | Мужские цветки в бутоне . | Мужские цветки открытые . | Опадание мужских цветков . | Женские цветки в бутоне . | Женские цветки открытые . | |
|---|---|---|---|---|---|---|
| 1 | 3 | — | — | 1 | — | |
| 2 | 2 | 1 | — | 1 | — | |
| 3 | 2 | — | 1 | 1 | — | |
| 4 | 1 | 1 | 1 | 1 | — | |
| 5 | — | 1 | 2 | 1 | — | |
| 6 | ||||||
| 6 | — | — | — | — | — | 1 |
Персонажи Обмениваются между мужскими и женскими цветами
На рисунках 3–7 показаны различные стадии развития, идентифицированные для мужских цветков (рис.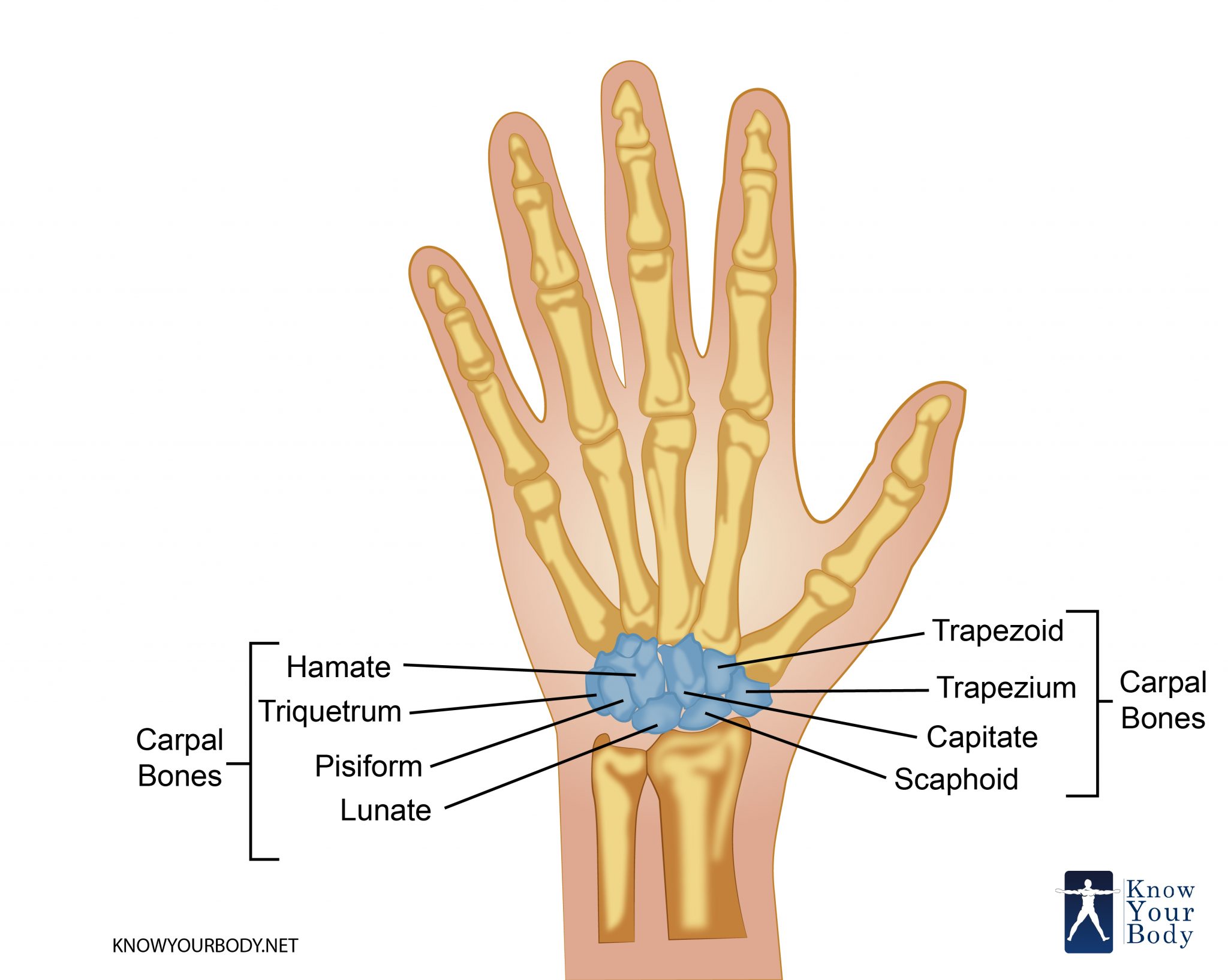 3 и 4) и женских цветков (рис. 5–7) G.аттенуата . Было замечено, что ряд признаков является общим для мужских и женских цветков. На поперечном срезе оба типа цветков были заметно треугольными вскоре после закладки, что особенно заметно у женских цветков (рис. 2B, C, 3E и 5B), хотя в конце развития они стали в основном округлыми. Хотя цветоножка была чрезвычайно короткой, а цветки слегка утопленными в рахиллах, можно было наблюдать заметное основание цветка высотой 0,5–1 мм (рис. 3D и 5A).Организация ткани у основания цветков была одинаковой для обоих полов на разных наблюдаемых стадиях развития. Как только чашелистики отделились друг от друга в этой области и лепестки начали отделяться, наблюдалась васкуляризация в виде внешнего кольца, состоящего из шести сосудистых пучков, и центрального сосудистого комплекса, образованного прокамбиальными тяжами (рис. 3E и 6C). Наружное кольцо четко соответствовало базальной васкуляризации стерильного или фертильного андроцея. Центральный комплекс объединил три уже дифференцированных сосудистых пучка, обслуживающих три доли фертильного или стерильного гинецея.
3 и 4) и женских цветков (рис. 5–7) G.аттенуата . Было замечено, что ряд признаков является общим для мужских и женских цветков. На поперечном срезе оба типа цветков были заметно треугольными вскоре после закладки, что особенно заметно у женских цветков (рис. 2B, C, 3E и 5B), хотя в конце развития они стали в основном округлыми. Хотя цветоножка была чрезвычайно короткой, а цветки слегка утопленными в рахиллах, можно было наблюдать заметное основание цветка высотой 0,5–1 мм (рис. 3D и 5A).Организация ткани у основания цветков была одинаковой для обоих полов на разных наблюдаемых стадиях развития. Как только чашелистики отделились друг от друга в этой области и лепестки начали отделяться, наблюдалась васкуляризация в виде внешнего кольца, состоящего из шести сосудистых пучков, и центрального сосудистого комплекса, образованного прокамбиальными тяжами (рис. 3E и 6C). Наружное кольцо четко соответствовало базальной васкуляризации стерильного или фертильного андроцея. Центральный комплекс объединил три уже дифференцированных сосудистых пучка, обслуживающих три доли фертильного или стерильного гинецея. Абаксиальный эпидермис всех органов околоцветника характеризовался наличием сплошного слоя сосочков, которые становились особенно заметными по мере развития (рис. 3G, 5B и 7G).
Абаксиальный эпидермис всех органов околоцветника характеризовался наличием сплошного слоя сосочков, которые становились особенно заметными по мере развития (рис. 3G, 5B и 7G).
Рис. 3.
Стадии развития мужского цветка Gaussia attenuata , I. (A) Мужской цветок в ранней бутоне (вид сверху на СЭМ, околоцветник удален). (B) Мужской цветок в поздней бутоне (СЭМ вид сбоку, околоцветник и две тычинки удалены). (C) Мужской цветок в поздней бутоне (вид сверху на СЭМ, околоцветник удален).(D) Продольный разрез мужского цветка в бутоне. (E) Поперечный разрез мужского цветка в ранней бутоне по направлению к базальной области. (F) Поперечный срез мужского цветка в ранней бутоне на уровне середины высоты пестика, демонстрирующий ранние развивающиеся материнские клетки нектарника и микроспоры. (G) Поперечный срез мужского цветка в поздней бутоне по направлению к основанию пестика. (H) Поперечный срез мужского цветка на поздней стадии бутона по направлению к середине высоты пестика, демонстрирующий деградацию тапетала и более позднюю стадию развития нектарника. (I) Поперечный срез, показывающий детали андроцея и пестика мужского цветка на поздней стадии бутонизации. Шкала баров: (А) = 250 мкм; (В) = 500 мкм; (С) = 500 мкм; (D) = 200 мкм; (Е) = 200 мкм; (F) = 50 мкм; (G) = 200 мкм; (Н) = 50 мкм; (I) = 100 мкм. Cvb, центральный сосудистый пучок; Et — эпителиальная ткань; Mi – материнские клетки микроспор; Пе, лепесток; Ptd, пестик; Pvb — пучок периферических сосудов; Se, чашелистик; ст, тычинка; Тп, тапетум.
(I) Поперечный срез, показывающий детали андроцея и пестика мужского цветка на поздней стадии бутонизации. Шкала баров: (А) = 250 мкм; (В) = 500 мкм; (С) = 500 мкм; (D) = 200 мкм; (Е) = 200 мкм; (F) = 50 мкм; (G) = 200 мкм; (Н) = 50 мкм; (I) = 100 мкм. Cvb, центральный сосудистый пучок; Et — эпителиальная ткань; Mi – материнские клетки микроспор; Пе, лепесток; Ptd, пестик; Pvb — пучок периферических сосудов; Se, чашелистик; ст, тычинка; Тп, тапетум.
Рис. 3.
Стадии развития мужского цветка Gaussia attenuata , I.(A) Мужской цветок в ранней бутоне (вид сверху на СЭМ, околоцветник удален). (B) Мужской цветок в поздней бутоне (СЭМ вид сбоку, околоцветник и две тычинки удалены). (C) Мужской цветок в поздней бутоне (вид сверху на СЭМ, околоцветник удален). (D) Продольный разрез мужского цветка в бутоне. (E) Поперечный разрез мужского цветка в ранней бутоне по направлению к базальной области. (F) Поперечный срез мужского цветка в ранней бутоне на уровне середины высоты пестика, демонстрирующий ранние развивающиеся материнские клетки нектарника и микроспоры. (G) Поперечный срез мужского цветка в поздней бутоне по направлению к основанию пестика. (H) Поперечный срез мужского цветка на поздней стадии бутона по направлению к середине высоты пестика, демонстрирующий деградацию тапетала и более позднюю стадию развития нектарника. (I) Поперечный срез, показывающий детали андроцея и пестика мужского цветка на поздней стадии бутонизации. Шкала баров: (А) = 250 мкм; (В) = 500 мкм; (С) = 500 мкм; (D) = 200 мкм; (Е) = 200 мкм; (F) = 50 мкм; (G) = 200 мкм; (Н) = 50 мкм; (I) = 100 мкм.Cvb, центральный сосудистый пучок; Et — эпителиальная ткань; Mi – материнские клетки микроспор; Пе, лепесток; Ptd, пестик; Pvb — пучок периферических сосудов; Se, чашелистик; ст, тычинка; Тп, тапетум.
(G) Поперечный срез мужского цветка в поздней бутоне по направлению к основанию пестика. (H) Поперечный срез мужского цветка на поздней стадии бутона по направлению к середине высоты пестика, демонстрирующий деградацию тапетала и более позднюю стадию развития нектарника. (I) Поперечный срез, показывающий детали андроцея и пестика мужского цветка на поздней стадии бутонизации. Шкала баров: (А) = 250 мкм; (В) = 500 мкм; (С) = 500 мкм; (D) = 200 мкм; (Е) = 200 мкм; (F) = 50 мкм; (G) = 200 мкм; (Н) = 50 мкм; (I) = 100 мкм.Cvb, центральный сосудистый пучок; Et — эпителиальная ткань; Mi – материнские клетки микроспор; Пе, лепесток; Ptd, пестик; Pvb — пучок периферических сосудов; Se, чашелистик; ст, тычинка; Тп, тапетум.
Рис. 4.
Стадии развития мужского цветка Gaussia attenuata , II. (A) Поперечный срез мужского цветка в поздней бутоне через базальную область пестика. (B) Поперечный срез, показывающий детали пестика на уровне нектарника, обнажая эпителиальную ткань (цветок в поздней почке). (C) Поперечный срез мужского цветка в поздней бутоне, показывающий астивацию створки (пестиллодия средней высоты). (D) Поперечный срез, показывающий детали пестика на средней высоте (цветок в поздней бутоне). (E) Поперечный срез на верхнем уровне мужского цветка в поздней бутоне, обнажающий верхушечные доли пестика. (F) Поперечный срез мужского цветка при цветении, показывающий базальную область пестика в центре и зрелые пыльцевые зерна. (G) Поперечный срез мужского цветка при цветении, показывающий детали пестика, показывающие позднюю стадию развития нектара и многочисленные шовные идиобласты.(H) Поперечный срез мужского цветка при цветении на уровне немного ниже вершины пестика. (I) Деталь пестика, показывающая сосудистую сеть и идиобласты шва. Шкала баров: (А) = 200 мкм; (В) = 100 мкм; (С) = 500 мкм; (D) = 100 мкм; (Е) = 100 мкм; (F) = 200 мкм; (G) = 100 мкм; (Н) = 500 мкм; (I) = 100 мкм. А, пыльник; Et — эпителиальная ткань; Пе, лепесток; Po, пыльцевые зерна; Ptd, пестик; Ri, шовный идиобласт, St, тычинка; Vb, сосудистый пучок.
(C) Поперечный срез мужского цветка в поздней бутоне, показывающий астивацию створки (пестиллодия средней высоты). (D) Поперечный срез, показывающий детали пестика на средней высоте (цветок в поздней бутоне). (E) Поперечный срез на верхнем уровне мужского цветка в поздней бутоне, обнажающий верхушечные доли пестика. (F) Поперечный срез мужского цветка при цветении, показывающий базальную область пестика в центре и зрелые пыльцевые зерна. (G) Поперечный срез мужского цветка при цветении, показывающий детали пестика, показывающие позднюю стадию развития нектара и многочисленные шовные идиобласты.(H) Поперечный срез мужского цветка при цветении на уровне немного ниже вершины пестика. (I) Деталь пестика, показывающая сосудистую сеть и идиобласты шва. Шкала баров: (А) = 200 мкм; (В) = 100 мкм; (С) = 500 мкм; (D) = 100 мкм; (Е) = 100 мкм; (F) = 200 мкм; (G) = 100 мкм; (Н) = 500 мкм; (I) = 100 мкм. А, пыльник; Et — эпителиальная ткань; Пе, лепесток; Po, пыльцевые зерна; Ptd, пестик; Ri, шовный идиобласт, St, тычинка; Vb, сосудистый пучок.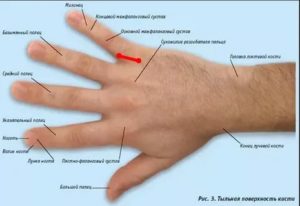
Рис. 4.
Стадии развития мужского цветка Gaussia attenuata , II.(A) Поперечный срез мужского цветка в поздней бутоне через базальную область пестика. (B) Поперечный срез, показывающий детали пестика на уровне нектарника, обнажая эпителиальную ткань (цветок в поздней почке). (C) Поперечный срез мужского цветка в поздней бутоне, показывающий астивацию створки (пестиллодия средней высоты). (D) Поперечный срез, показывающий детали пестика на средней высоте (цветок в поздней бутоне). (E) Поперечный срез на верхнем уровне мужского цветка в поздней бутоне, обнажающий верхушечные доли пестика.(F) Поперечный срез мужского цветка при цветении, показывающий базальную область пестика в центре и зрелые пыльцевые зерна. (G) Поперечный срез мужского цветка при цветении, показывающий детали пестика, показывающие позднюю стадию развития нектара и многочисленные шовные идиобласты. (H) Поперечный срез мужского цветка при цветении на уровне немного ниже вершины пестика. (I) Деталь пестика, показывающая сосудистую сеть и идиобласты шва.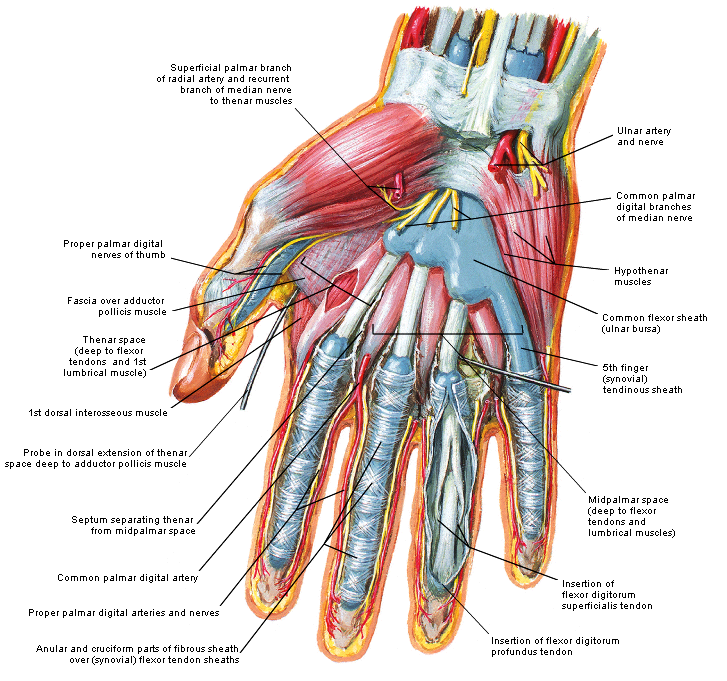 Шкала баров: (А) = 200 мкм; (В) = 100 мкм; (С) = 500 мкм; (D) = 100 мкм; (Е) = 100 мкм; (F) = 200 мкм; (G) = 100 мкм; (Н) = 500 мкм; (I) = 100 мкм.А, пыльник; Et — эпителиальная ткань; Пе, лепесток; Po, пыльцевые зерна; Ptd, пестик; Ri, шовный идиобласт, St, тычинка; Vb, сосудистый пучок.
Шкала баров: (А) = 200 мкм; (В) = 100 мкм; (С) = 500 мкм; (D) = 100 мкм; (Е) = 100 мкм; (F) = 200 мкм; (G) = 100 мкм; (Н) = 500 мкм; (I) = 100 мкм.А, пыльник; Et — эпителиальная ткань; Пе, лепесток; Po, пыльцевые зерна; Ptd, пестик; Ri, шовный идиобласт, St, тычинка; Vb, сосудистый пучок.
Рис. 5.
Стадии развития женского цветка Gaussia attenuata , I. (A) Продольный разрез женского цветка в ранней бутоне, показывающий развивающийся стаминодий. (B) Поперечный срез женского цветка в начале бутона на средней высоте. (C) Фрагмент гинецея в поперечном срезе на стадии ранней цветочной почки, на котором видны масляные тела в венчике.(D) СЭМ гинецея и стерильного андроцея ранней цветочной почки (удален околоцветник). (E) СЭМ гинецея и стерильного андроцея на несколько более поздней стадии (удален околоцветник). Шкала баров: (А) = 200 мкм; (В) = 200 мкм; (С) = 100 мкм; (D) = 100 мкм; (Е) = 50 мкм. Са, плодолистик; Cp, центральный выступ; Пе, лепесток; Se, чашелистик; Std, стаминод.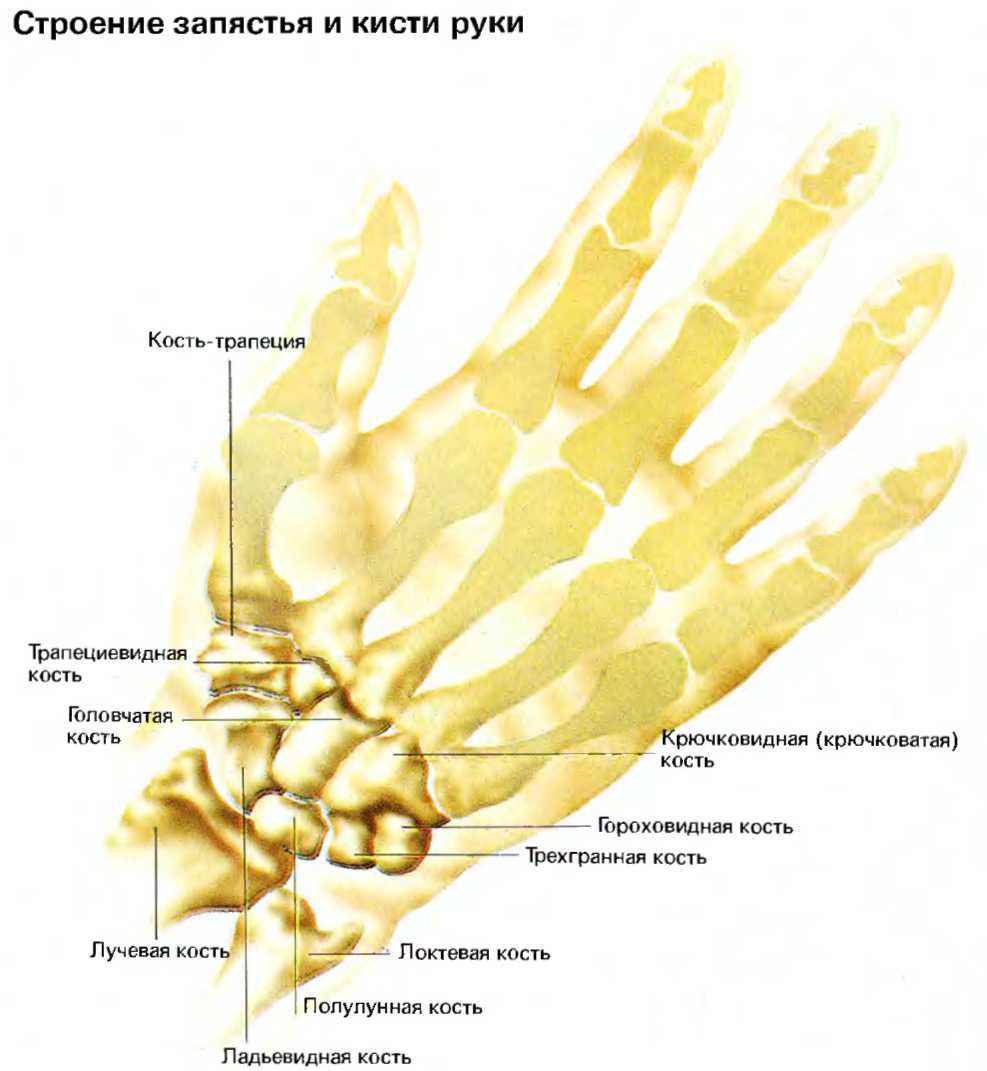
Рис. 5.
Стадии развития женского цветка Gaussia attenuata , I. (A) Продольный разрез женского цветка в ранней бутоне, показывающий развивающийся стаминодий.(B) Поперечный срез женского цветка в начале бутона на средней высоте. (C) Фрагмент гинецея в поперечном срезе на стадии ранней цветочной почки, на котором видны масляные тела в венчике. (D) СЭМ гинецея и стерильного андроцея ранней цветочной почки (удален околоцветник). (E) СЭМ гинецея и стерильного андроцея на несколько более поздней стадии (удален околоцветник). Шкала баров: (А) = 200 мкм; (В) = 200 мкм; (С) = 100 мкм; (D) = 100 мкм; (Е) = 50 мкм. Са, плодолистик; Cp, центральный выступ; Пе, лепесток; Se, чашелистик; Std, стаминод.
Рис. 6.
Стадии развития женского цветка Gaussia attenuata , II. (A) SEM вид сверху на гинецей (удален околоцветник). (B) SEM боковой вид гинецея (удален околоцветник). (C) Поперечный срез на уровне основания цветка. (D) Поперечный срез цветка на уровне локул.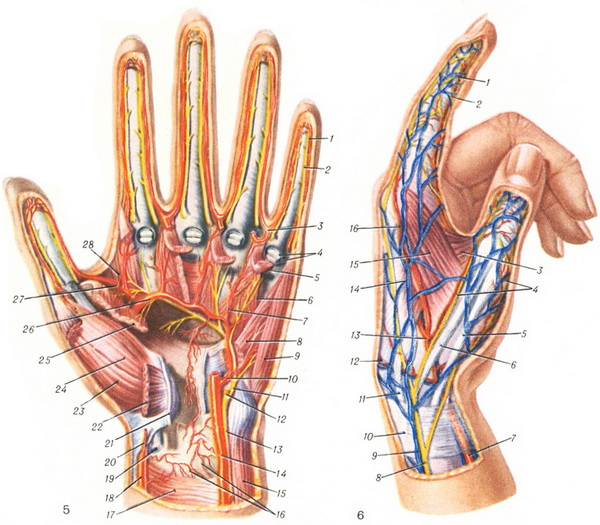 (E) Деталь глазной области, показывающая развивающийся септальный нектарник. (F) Поперечный срез цветка на средней высоте столбика, обнажающий шовные идиобласты, скопившиеся в стенках плодолистика.(G) Поперечный срез на средней высоте стиля, показывающий васкуляризацию плодолистиков и передающего тракта пыльцевой трубки. (H) Поперечный срез на верхнем уровне стиля, показывающий компитум. (I) Поперечный срез на верхнем уровне цветка, обнажающий рыльце рыльца. Шкала баров: (А) = 250 мкм; (В) = 250 мкм; (С) = 500 мкм; (D) = 200 мкм; (Е) = 100 мкм; (F) = 500 мкм; (G) = 100 мкм; (Н) = 100 мкм; (I) = 200 мкм. Са, плодолистик; Ср, компитум; Et — эпителиальная ткань; Гр, гинецей; Ов, семязачаток; Пе, лепесток; Pt — пыльцевые трубки, передающие тракты; Ri — шовный идиобласт; Se, чашелистик; Sl, стигматическая доля; Std, стаминодия; Vb, сосудистый пучок.
(E) Деталь глазной области, показывающая развивающийся септальный нектарник. (F) Поперечный срез цветка на средней высоте столбика, обнажающий шовные идиобласты, скопившиеся в стенках плодолистика.(G) Поперечный срез на средней высоте стиля, показывающий васкуляризацию плодолистиков и передающего тракта пыльцевой трубки. (H) Поперечный срез на верхнем уровне стиля, показывающий компитум. (I) Поперечный срез на верхнем уровне цветка, обнажающий рыльце рыльца. Шкала баров: (А) = 250 мкм; (В) = 250 мкм; (С) = 500 мкм; (D) = 200 мкм; (Е) = 100 мкм; (F) = 500 мкм; (G) = 100 мкм; (Н) = 100 мкм; (I) = 200 мкм. Са, плодолистик; Ср, компитум; Et — эпителиальная ткань; Гр, гинецей; Ов, семязачаток; Пе, лепесток; Pt — пыльцевые трубки, передающие тракты; Ri — шовный идиобласт; Se, чашелистик; Sl, стигматическая доля; Std, стаминодия; Vb, сосудистый пучок.
Рис. 6.
Стадии развития женского цветка Gaussia attenuata , II. (A) SEM вид сверху на гинецей (удален околоцветник). (B) SEM боковой вид гинецея (удален околоцветник). (C) Поперечный срез на уровне основания цветка. (D) Поперечный срез цветка на уровне локул. (E) Деталь глазной области, показывающая развивающийся септальный нектарник. (F) Поперечный срез цветка на средней высоте столбика, обнажающий шовные идиобласты, скопившиеся в стенках плодолистика.(G) Поперечный срез на средней высоте стиля, показывающий васкуляризацию плодолистиков и передающего тракта пыльцевой трубки. (H) Поперечный срез на верхнем уровне стиля, показывающий компитум. (I) Поперечный срез на верхнем уровне цветка, обнажающий рыльце рыльца. Шкала баров: (А) = 250 мкм; (В) = 250 мкм; (С) = 500 мкм; (D) = 200 мкм; (Е) = 100 мкм; (F) = 500 мкм; (G) = 100 мкм; (Н) = 100 мкм; (I) = 200 мкм. Са, плодолистик; Ср, компитум; Et — эпителиальная ткань; Гр, гинецей; Ов, семязачаток; Пе, лепесток; Pt — пыльцевые трубки, передающие тракты; Ri — шовный идиобласт; Se, чашелистик; Sl, стигматическая доля; Std, стаминодия; Vb, сосудистый пучок.
(B) SEM боковой вид гинецея (удален околоцветник). (C) Поперечный срез на уровне основания цветка. (D) Поперечный срез цветка на уровне локул. (E) Деталь глазной области, показывающая развивающийся септальный нектарник. (F) Поперечный срез цветка на средней высоте столбика, обнажающий шовные идиобласты, скопившиеся в стенках плодолистика.(G) Поперечный срез на средней высоте стиля, показывающий васкуляризацию плодолистиков и передающего тракта пыльцевой трубки. (H) Поперечный срез на верхнем уровне стиля, показывающий компитум. (I) Поперечный срез на верхнем уровне цветка, обнажающий рыльце рыльца. Шкала баров: (А) = 250 мкм; (В) = 250 мкм; (С) = 500 мкм; (D) = 200 мкм; (Е) = 100 мкм; (F) = 500 мкм; (G) = 100 мкм; (Н) = 100 мкм; (I) = 200 мкм. Са, плодолистик; Ср, компитум; Et — эпителиальная ткань; Гр, гинецей; Ов, семязачаток; Пе, лепесток; Pt — пыльцевые трубки, передающие тракты; Ri — шовный идиобласт; Se, чашелистик; Sl, стигматическая доля; Std, стаминодия; Vb, сосудистый пучок.
Рис.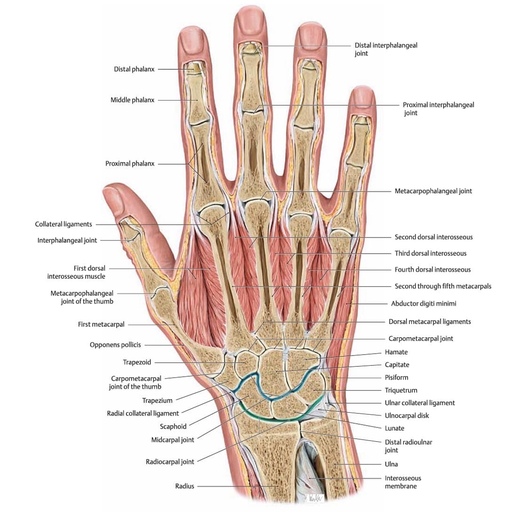 7.
7.
Стадии развития женского цветка Gaussia attenuata , III (антезис). (A) SEM боковой вид гинецея (удален околоцветник). (B) SEM боковой вид стигмы, показывающий внутреннюю поверхность сосочков. (C) Продольный разрез цветка по направлению к центру гинецея, открывающий нектарные отверстия. (D) Поперечный срез цветка на верхнем уровне локул. (E) Деталь перегородочного нектарника в поперечном сечении, показывающая заметную эпителиальную ткань и отверстие нектарника.(F) Продольный срез очага, показывающий эпителиальную ткань, прилегающую к канатику, микропиле и слой танниновых идиобластов, окружающих нуцеллус. (G) Поперечный срез лепестка, показывающий стегмы, связанные с сосудистой тканью и сосочковым абаксиальным эпидермисом. Шкала баров: (А) = 500 мкм; (В) = 100 мкм; (С) = 200 мкм; (D) = 500 мкм; (Е) = 100 мкм; (F) = 100 мкм; (G) = 50 мкм. Са, плодолистик; Std, стаминодия; Sl, стигматическая доля; Пе, лепесток; Pt — пыльцевые трубки, передающие тракты; Ов, семязачаток; Op, открытие нектара; Гр, гинецей; Et — эпителиальная ткань; Mi, микропиле; Ti – дубильный идиобласт; Stg, стегмы; Па, папилла.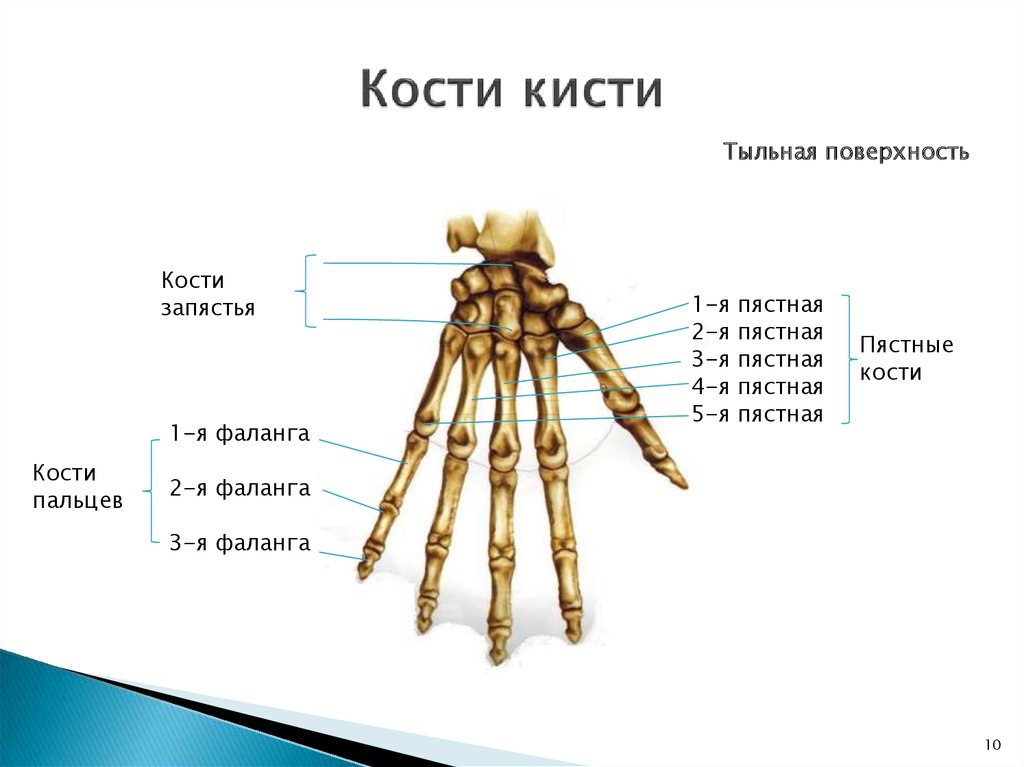
Рис. 7.
Стадии развития женского цветка Gaussia attenuata , III (антезис). (A) SEM боковой вид гинецея (удален околоцветник). (B) SEM боковой вид стигмы, показывающий внутреннюю поверхность сосочков. (C) Продольный разрез цветка по направлению к центру гинецея, открывающий нектарные отверстия. (D) Поперечный срез цветка на верхнем уровне локул. (E) Деталь перегородочного нектарника в поперечном сечении, показывающая заметную эпителиальную ткань и отверстие нектарника.(F) Продольный срез очага, показывающий эпителиальную ткань, прилегающую к канатику, микропиле и слой танниновых идиобластов, окружающих нуцеллус. (G) Поперечный срез лепестка, показывающий стегмы, связанные с сосудистой тканью и сосочковым абаксиальным эпидермисом. Шкала баров: (А) = 500 мкм; (В) = 100 мкм; (С) = 200 мкм; (D) = 500 мкм; (Е) = 100 мкм; (F) = 100 мкм; (G) = 50 мкм. Са, плодолистик; Std, стаминодия; Sl, стигматическая доля; Пе, лепесток; Pt — пыльцевые трубки, передающие тракты; Ов, семязачаток; Op, открытие нектара; Гр, гинецей; Et — эпителиальная ткань; Mi, микропиле; Ti – дубильный идиобласт; Stg, стегмы; Па, папилла.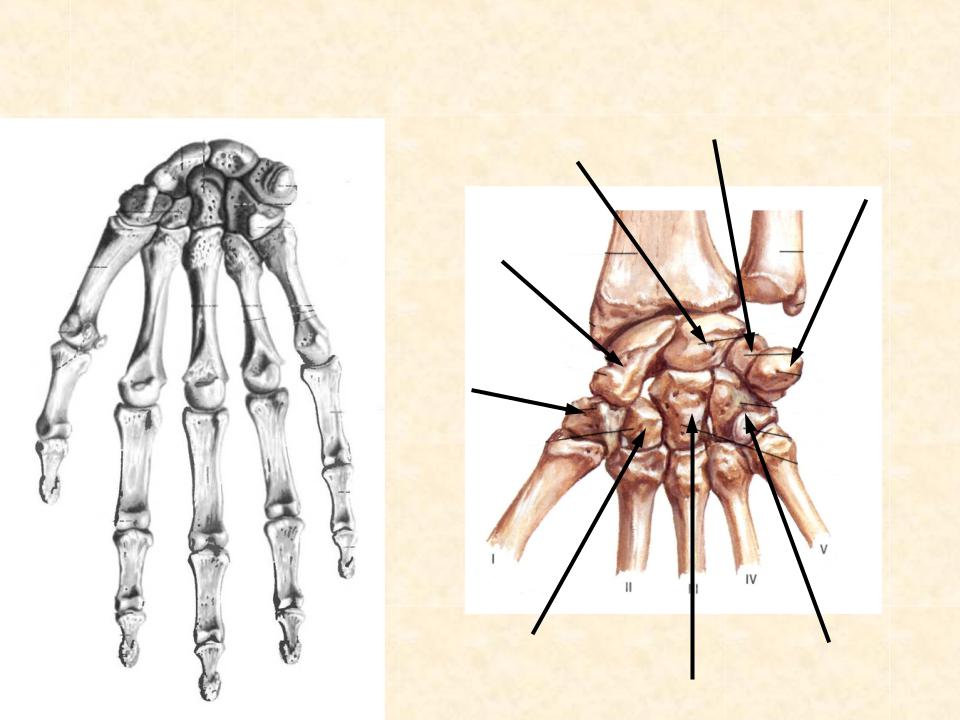
Строение и развитие мужского цветка
Морфология
Чашечка образована тремя свободными черепитчатыми чашелистиками (рис. 3E, G). Они были вогнутыми, округлыми и заметно мясистыми от основания до середины высоты, тогда как к вершине и краям они были тонкими (рис. 3D). В ранней зародыше они были длиной ∼0,5–1 мм и шириной 1 мм и полностью покрывали внутренние органы (рис. 2С). Однако по мере роста внутренних органов чашелистики стали постепенно раскрываться, становясь клиновидными и немного истончаясь, достигая к зрелости около 2 мм в ширину и 1–1,5 мм в длину (рис.2Д, Е).
Венчик состоял из трех створчатых лепестков (рис. 2F, G). Они были вогнутыми, мясистыми по всей длине и в ограниченной степени срастались с андроцеем у основания (рис. 3D). На ранних стадиях развития, когда чашелистики уже отделились и стали видны лепестки, последние имели длину ~1–1,5 мм и диаметр 1 мм (рис. 2В, Е). Лепестки плотно охватывают репродуктивные органы, чему способствуют эпидермальные сосочки, которые на боковых краях действовали как застежка-молния (рис.2Г). Как только венчик открылся, позволив внутренним органам расшириться, лепестки начали отгибаться, достигая ~2–2,5 мм в длину и 1,5–2 мм в ширину при цветении (рис. 2G, J).
2В, Е). Лепестки плотно охватывают репродуктивные органы, чему способствуют эпидермальные сосочки, которые на боковых краях действовали как застежка-молния (рис.2Г). Как только венчик открылся, позволив внутренним органам расшириться, лепестки начали отгибаться, достигая ~2–2,5 мм в длину и 1,5–2 мм в ширину при цветении (рис. 2G, J).
Андроцей состоял из шести тычинок, организованных в два оборота по три тычинки, одна напротив чашелистиков, а другая против лепестков (рис. 3A–C, G). Все они были фертильными, прикреплялись на одном уровне и слегка срастались с венчиком у основания (рис. 3D). На самой ранней изученной стадии развития (рис.3A, D) видно, что пыльники уже были двухлопастными, и каждая доля начинала расщепляться на два разных гнезда. К этому времени пыльники имели ширину ~0,2 мм и длину 0,3–0,4 мм, а их форма частично определялась взаимодействием с другими тесно прижатыми пыльниками и пестиком. Нити были различимы, хотя некоторые из них еще не начали удлиняться; большинство из них были очень короткими с максимальной длиной 0,1 мм (рис.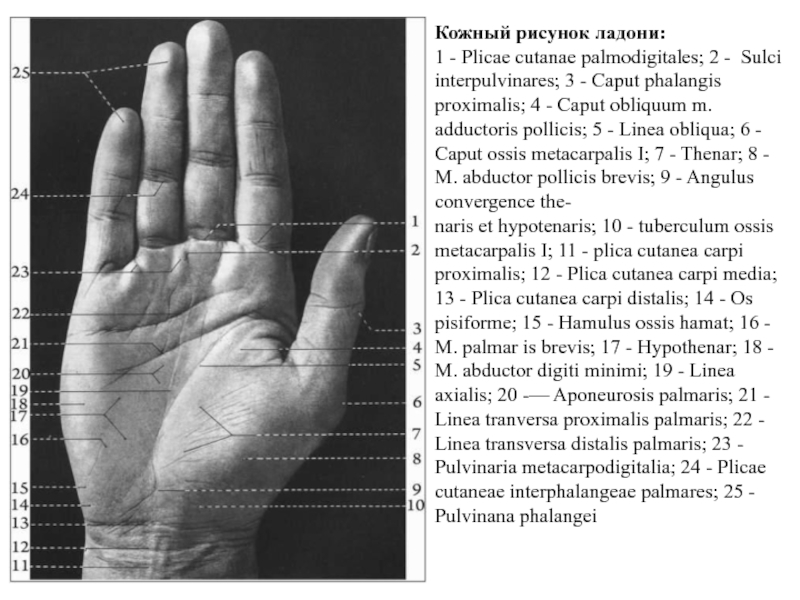 3D). На более позднем этапе развития и пыльники, и нити одновременно удлиняются и отделяются от стерильного гинецея, достигая длины 1–1,5 мм непосредственно перед цветением и раскрытием венчика (рис.3Б, С). На этой стадии и андроцей, и пестик достигают одинаковой высоты. Примерно во время цветения лепестки отделялись вдоль своих боковых краев, позволяя нитям удлиняться, пока они не достигли длины ~ 1,5–2 мм (рис. 2G-J). В зрелых цветках нити имеют цилиндрическую и загнутую форму. На этом этапе пыльники были зафиксированы дорсально и демонстрировали внешнее расхождение через продольное стомиум, достигая ~ 2 мм в длину (рис. 2G–J и 4H).
3D). На более позднем этапе развития и пыльники, и нити одновременно удлиняются и отделяются от стерильного гинецея, достигая длины 1–1,5 мм непосредственно перед цветением и раскрытием венчика (рис.3Б, С). На этой стадии и андроцей, и пестик достигают одинаковой высоты. Примерно во время цветения лепестки отделялись вдоль своих боковых краев, позволяя нитям удлиняться, пока они не достигли длины ~ 1,5–2 мм (рис. 2G-J). В зрелых цветках нити имеют цилиндрическую и загнутую форму. На этом этапе пыльники были зафиксированы дорсально и демонстрировали внешнее расхождение через продольное стомиум, достигая ~ 2 мм в длину (рис. 2G–J и 4H).
В центре цветка видна очень заметная пестичная ножка.На самых ранних изученных стадиях развития пестик представлял собой компактный, более или менее треугольный столбик с двумя апикальными лопастями (рис. 3A–D), который в некоторых случаях становился апикально трехраздельным по мере приближения цветения (рис. 2J и 3B). Тем не менее, у большинства цветков, наблюдаемых в поздней бутоне или при цветении, можно было увидеть раздвоенный на вершине пестик (рис. 4H, I). Форма внешней поверхности определялась взаимодействием с внутренними областями близко расположенных пыльников (рис. 4А–Д). На ранних стадиях развития он имел длину 0,5 мм, достигая к зрелости ~2–2,5 мм.
4H, I). Форма внешней поверхности определялась взаимодействием с внутренними областями близко расположенных пыльников (рис. 4А–Д). На ранних стадиях развития он имел длину 0,5 мм, достигая к зрелости ~2–2,5 мм.
Анатомия
Три сосудистых пучка, два латеральных и один центральный, наблюдались у основания каждого чашелистика, ориентированного к адаксиальной поверхности (рис. 3E, G). Боковые пучки исчезли на середине длины чашелистика, и только центральный пучок протянулся несколько дальше вверх, отсутствуя к вершине (рис. 3G). В лепестках было от пяти до девяти сосудистых пучков в базальной области и от трех до пяти к вершине (рис. 3G и 4C, E, H).Характер васкуляризации околоцветника оставался постоянным по мере развития.
Один сосудистый пучок омывал каждый филамент по всей его длине, достигая соединительной области (рис. 3I и 4A, E, H). Этот пучок наблюдался с самых ранних изученных стадий развития (таблица 1), когда спорогенная ткань уже была дифференцирована (рис.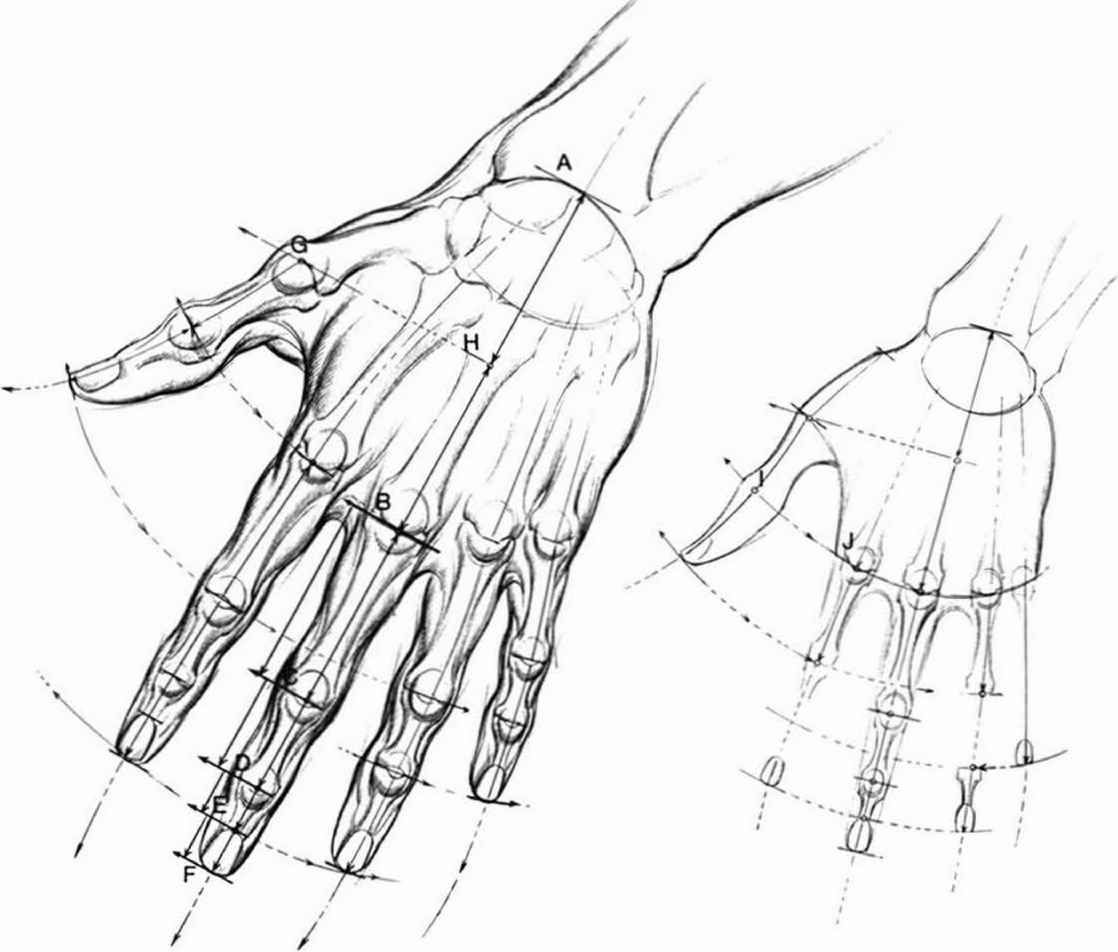 3F). На этой стадии мы наблюдали эпидермис пыльника, эндотеций, один средний слой и тапетум. На более позднем этапе развития тапетум распадался параллельно с образованием тетрады (рис.3Н, 1). На последних наблюдаемых стадиях эпидермис пыльника все еще присутствовал, эндотеций развил волокнистые утолщения стенок, а пыльцевые зерна были зрелыми (рис. 4F, H).
3F). На этой стадии мы наблюдали эпидермис пыльника, эндотеций, один средний слой и тапетум. На более позднем этапе развития тапетум распадался параллельно с образованием тетрады (рис.3Н, 1). На последних наблюдаемых стадиях эпидермис пыльника все еще присутствовал, эндотеций развил волокнистые утолщения стенок, а пыльцевые зерна были зрелыми (рис. 4F, H).
Помимо того, что сосудистые тяжи были лучше выражены в зрелых цветках, в сосудистой сети пестика не наблюдалось существенных изменений в процессе его развития. Было замечено, что три сосудистых пучка проходят через центр пестика от базального уровня до середины высоты, простираясь немного ниже вершины (рис.4Г, Ж, И), хотя на самой вершине их не наблюдалось (не показано). У двухлопастного пестика большая доля обычно обслуживается двумя сосудистыми пучками (рис. 4G, I). На средней высоте во внутренних областях пестика были обнаружены эпителиальные клетки, которые образовывали нектароподобную структуру, состоящую из независимых каналов (рис. 4G). Видно, что развитие нектарника начинается в ранних цветочных почках, в промежутках между лопастями пестика (рис. 3F, H). Однако по мере развития эпителиальные клетки дифференцируются в четко очерченную извитую секреторную ткань, которая сообщается с внешней поверхностью через боковые отверстия (рис.4Б, Г).
4G). Видно, что развитие нектарника начинается в ранних цветочных почках, в промежутках между лопастями пестика (рис. 3F, H). Однако по мере развития эпителиальные клетки дифференцируются в четко очерченную извитую секреторную ткань, которая сообщается с внешней поверхностью через боковые отверстия (рис.4Б, Г).
Дубильные идиобласты в основании цветка наблюдались только вблизи эпидермиса (рис. 3E). В органах околоцветника они были разбросаны по всему мезофиллу, в основном по направлению к абаксиальному эпидермису, будучи более многочисленными к основанию (рис. 3E), картина, которая оставалась постоянной по мере развития.
Предполагаемые масляные тела разных размеров наблюдались преимущественно в репродуктивных органах. В пределах андроцея они были плотно сгруппированы в эндотеции (преимущественно к середине высоты пыльника), эпидермальных клетках пыльника, соединительных тканях и вблизи стомиума (рис.3И). Они образовали заметное кольцо, окружающее сосудистый пучок каждой нити. Толщина этого кольца могла достигать трех-четырех клеточных слоев к середине высоты и в основании нитей. Масляные тельца также присутствовали в пестике, связанные с эпителиальными тканями нектарника (рис. 3F и 4B), тогда как к основанию или к дистальному концу их стало мало. Обилие масляных телец было значительно выше в цветках на ранних стадиях развития, чем в цветках в поздней бутонизации или в стадии цветения, на которых они наблюдались редко.
Толщина этого кольца могла достигать трех-четырех клеточных слоев к середине высоты и в основании нитей. Масляные тельца также присутствовали в пестике, связанные с эпителиальными тканями нектарника (рис. 3F и 4B), тогда как к основанию или к дистальному концу их стало мало. Обилие масляных телец было значительно выше в цветках на ранних стадиях развития, чем в цветках в поздней бутонизации или в стадии цветения, на которых они наблюдались редко.
Идиобласты, содержащие рафид, в прикорневой области цветка не обнаружены. Однако было замечено, что они были обильно распределены в мезофилле, в основном в нижней центральной части околоцветника, на протяжении всего его развития (рис. 3G и 4A, E). Рафидные идиобласты также наблюдались в андроцее, ограниченном тапетумом цветков возле цветения (рис. 4A, C, E). Они также были замечены в пестиках, становясь более заметными по мере продвижения к цветению, хотя сначала они появлялись в цветочных почках на более ранней стадии, всегда ограничиваясь периферией (рис. 4Б, Д, Ж, И).
4Б, Д, Ж, И).
Строение и развитие женского цветка
Морфология
Чашечка женского цветка была образована тремя черепитчатыми мясистыми чашелистиками, которые были толще чашелистиков мужского цветка (рис. 5А, В). В начале развития перекрывающиеся чашелистики полностью окружали внутренние цветочные органы, придавая цветку характерную треугольную форму (рис. 2B, C и 5B). На этой стадии чашелистики были сильно вогнутыми, достигая ∼0,5 мм в длину и ∼1 мм в ширину, тогда как к цветению их размер удваивался до ∼1 мм в длину и ∼2 мм в ширину, их края слегка загибались к верхушке. вершина (рис.6С, Г).
Венчик женских цветков состоял из трех лепестков, почти полностью клапанных, но слегка черепитчатых к основанию (рис. 6C). В ранних цветочных почках они были более или менее треугольными и вогнутыми, ∼1–1,5 мм в длину и ∼1 мм в ширину, герметично охватывая внутренние мутовки (рис. 2F, G). Тем не менее, при цветении венчик раскрывался, и лопасти лепестков сильно загибались, достигая 2 мм в длину и 2 мм в ширину к основанию (рис. 2Л, М).
2F, G). Тем не менее, при цветении венчик раскрывался, и лопасти лепестков сильно загибались, достигая 2 мм в длину и 2 мм в ширину к основанию (рис. 2Л, М).
Стерильный андроцей состоял из шести стаминодий.Инициация развития этих органов происходила в два этапа, на каждом из которых образовывался отдельный оборот (рис. 5D, E). Первый сформированный оборот (внешний оборот) состоял из трех антесепальных стаминодий (рис. 7А). Второй (внутренний) оборот состоял из трех переднелепестных стаминодий, которые также в основании срастались с плодолистиками (рис. 6C и 7A, C). На самых ранних наблюдаемых стадиях (рис. 5D, E) органы наружного оборота были немного крупнее органов внутреннего оборота.В это время стаминодии появились в виде очень коротких вогнутых уплощенных выростов. Впоследствии было замечено, что стаминодии продолжают свое удлинение, демонстрируя цилиндрическую, шиловидную, нитевидную форму по мере достижения цветения (рис. 6B и 7A, C). Несмотря на то, что стаминодии при цветении достигали длины почти 1 мм, образования дифференцированного пыльника не наблюдалось (рис. 2L и 7A).
2L и 7A).
Гинецей образован тремя плодолистиками, все из которых фертильны и расположены напротив чашелистиков.Самая ранняя наблюдаемая стадия развития плодолистика представляла собой три свободных очень коротких клиновидных выступа, образующих симметричный треугольник с заметным центральным выступом (рис. 5C, D). Впоследствии было замечено, что плодолистики претерпевают постгенитальное слияние через их боковые края, начиная с основания, с сопутствующим появлением вентральных щелей (рис. 5E). На более позднем этапе развития по мере их удлинения происходило соединение между боковыми сторонами соседних плодолистиков, что позволяло плодолистикам принимать свою характерную урцеолатную форму по мере приближения цветения (рис. 6A, B и 7A, C).В зрелом возрасте каждый плодолистик имел длину ~1,5 мм, асцидиальный у основания и складчатый к дистальному концу. Вместе три плодолистика составляли гинецей с базальной областью синаскидиата (рис. 6D и 7D) и дистальным симпликатным участком (рис. 6F, G), при этом плодолистики были свободны только к стигматическому концу (рис. 6I и 7A, B). Три самостоятельные складчатые доли, наблюдаемые в стигматической области, густо покрытые одноклеточными сосочками, консолидировались лишь непосредственно перед цветением, когда они также рефлекторировали (рис.7А, Б).
6F, G), при этом плодолистики были свободны только к стигматическому концу (рис. 6I и 7A, B). Три самостоятельные складчатые доли, наблюдаемые в стигматической области, густо покрытые одноклеточными сосочками, консолидировались лишь непосредственно перед цветением, когда они также рефлекторировали (рис.7А, Б).
Анатомия
В основании было видно, что чашелистики васкуляризированы с ранних стадий развития от одного до трех сосудистых пучков (рис. 5B), которые также наблюдались на средней длине и на поздних стадиях. Однако чашелистики не были васкуляризованы в их апикальной области ни на одной стадии развития. В лепестках в начале развития наблюдалось меньшее количество нитей: три были видны у основания цветка (рис. 5C), одна достигала вершины. На более позднем этапе развития на базальном уровне наблюдалось шесть или семь сосудистых пучков, на средней высоте пять и два или три в апикальной области (рис.6D, F, I). В основании каждого стаминодия на протяжении всего развития цветка можно было различить по одному сосудистому пучку.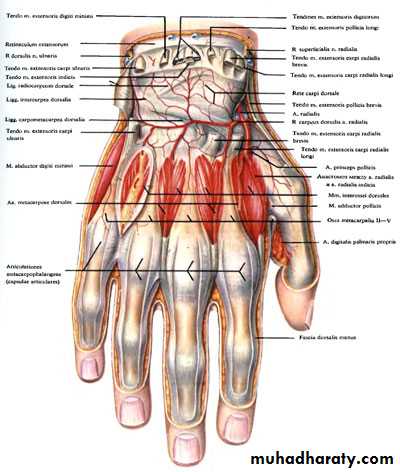 В плодолистиках по мере развития наблюдались поразительные изменения в васкуляризации. На ранних стадиях было замечено, что только одна сосудистая нить впадает в основание каждого плодолистика (рис. 5А). Параллельно с дифференцировкой плодолистиков наблюдалось разветвление сосудистых тяжей с образованием комплекса по крайней мере из одного дорсального и двух латеральных сосудистых пучков, орошающих каждый плодолистик из базальной области через среднюю область локул (рис.6D) и по всему стилю (рис. 6G). Тем не менее, васкуляризации, питающей стигматическую область, не наблюдалось.
В плодолистиках по мере развития наблюдались поразительные изменения в васкуляризации. На ранних стадиях было замечено, что только одна сосудистая нить впадает в основание каждого плодолистика (рис. 5А). Параллельно с дифференцировкой плодолистиков наблюдалось разветвление сосудистых тяжей с образованием комплекса по крайней мере из одного дорсального и двух латеральных сосудистых пучков, орошающих каждый плодолистик из базальной области через среднюю область локул (рис.6D) и по всему стилю (рис. 6G). Тем не менее, васкуляризации, питающей стигматическую область, не наблюдалось.
Верхняя часть столбика характеризовалась compitum, образованным между частично сросшимися дорсальными поверхностями плодолистиков и их сливающимися вентральными щелями (рис. 6H). Средняя часть столбика имеет треугольную форму в поперечном сечении, три плодолистика полностью срослись, а три вентральные щели остаются независимыми до точки, где они достигают локул (рис.6Е, Ж). Было замечено, что тракт передачи пыльцевой трубки (PTTT) является поверхностным, образованным слоем толщиной в одну клетку (рис. 6G), выстилающим апикальную складчатую зону плодолистиков и асцидиатную зону, где вентральные щели были независимыми. Было замечено, что ЧТТВ является секреторным по всей его длине (рис. 7C), включая локулу, где можно было наблюдать слой эпителиальных клеток, прилегающих к канатику (рис. 7F). В синаскидиальной области гинецея наблюдалось образование перегородочного нектарника, простирающегося вверх по всей длине локул.Было замечено, что он развивается из открытых каналов, образованных между соседними плодолистиками в глазковой области, как следствие их дифференциального постгенитального слияния (рис. 6D, E и 7E). Было замечено, что эти каналы сливаются по направлению к середине яичника и имеют открывающиеся протоки по направлению к внешнему эпидермису плодолистиков, от области средней высоты локул до их основания (рис. 7C–E). Было замечено, что нектарник состоит из эпителиальной секреторной ткани толщиной в один или, возможно, два клеточных слоя, образующих непрерывный слой от центральных каналов до трех наружных отверстий (рис.
6G), выстилающим апикальную складчатую зону плодолистиков и асцидиатную зону, где вентральные щели были независимыми. Было замечено, что ЧТТВ является секреторным по всей его длине (рис. 7C), включая локулу, где можно было наблюдать слой эпителиальных клеток, прилегающих к канатику (рис. 7F). В синаскидиальной области гинецея наблюдалось образование перегородочного нектарника, простирающегося вверх по всей длине локул.Было замечено, что он развивается из открытых каналов, образованных между соседними плодолистиками в глазковой области, как следствие их дифференциального постгенитального слияния (рис. 6D, E и 7E). Было замечено, что эти каналы сливаются по направлению к середине яичника и имеют открывающиеся протоки по направлению к внешнему эпидермису плодолистиков, от области средней высоты локул до их основания (рис. 7C–E). Было замечено, что нектарник состоит из эпителиальной секреторной ткани толщиной в один или, возможно, два клеточных слоя, образующих непрерывный слой от центральных каналов до трех наружных отверстий (рис. 7Е). Его можно рассматривать как промежуточное звено между сливно-простым и сливно-извитым типом нектарника. Было замечено, что каждый плодолистик имеет анатропную семяпочку, прикрепленную к гнезду на середине длины. Семяпочка была крассинуцеллятной, заполняя около 80 % глазной полости (рис. 7F). Наблюдалась папиллярная ткань, ограниченная зоной, прилегающей к канатику, но в основном связанная со стенкой глазного яблока, представляющая собой продолжение ЧЧТВ. Битегмическая семяпочка обладала наружным покровом, состоящим из десяти клеточных слоев толщиной посередине длины и около пяти клеточных слоев к концу микропиле.Внутренний покров был толщиной в два слоя клеток на всем протяжении. Микропиле формировалось исключительно внутренним покровом, который в этом месте плотно контактировал с сосочковой зоной глазной полости (рис. 7F).
7Е). Его можно рассматривать как промежуточное звено между сливно-простым и сливно-извитым типом нектарника. Было замечено, что каждый плодолистик имеет анатропную семяпочку, прикрепленную к гнезду на середине длины. Семяпочка была крассинуцеллятной, заполняя около 80 % глазной полости (рис. 7F). Наблюдалась папиллярная ткань, ограниченная зоной, прилегающей к канатику, но в основном связанная со стенкой глазного яблока, представляющая собой продолжение ЧЧТВ. Битегмическая семяпочка обладала наружным покровом, состоящим из десяти клеточных слоев толщиной посередине длины и около пяти клеточных слоев к концу микропиле.Внутренний покров был толщиной в два слоя клеток на всем протяжении. Микропиле формировалось исключительно внутренним покровом, который в этом месте плотно контактировал с сосочковой зоной глазной полости (рис. 7F).
Дубильные идиобласты можно было наблюдать рассеянными в мезофилле чашелистиков на протяжении всего их развития, в основном связанными с абаксиальной стороной (рис. 5А). Также видно, что нуцеллус окружает непрерывный двухклеточный слой, содержащий дубильные вещества (рис.7F). Масляные тела наблюдались во всех органах цветка, кроме стаминодий, от стадии ранней бутонизации до цветения. Было замечено, что они накапливаются в основании чашелистиков на всех стадиях развития, но распределяются по лепесткам на очень ранних стадиях развития (рис. 5А–С). По мере развития их становилось все больше в гинецее, особенно в базальной области, по направлению к внешней поверхности плодолистиков, а также в нектарных и стигматических сосочках (рис. 6E и 7E, F).
5А). Также видно, что нуцеллус окружает непрерывный двухклеточный слой, содержащий дубильные вещества (рис.7F). Масляные тела наблюдались во всех органах цветка, кроме стаминодий, от стадии ранней бутонизации до цветения. Было замечено, что они накапливаются в основании чашелистиков на всех стадиях развития, но распределяются по лепесткам на очень ранних стадиях развития (рис. 5А–С). По мере развития их становилось все больше в гинецее, особенно в базальной области, по направлению к внешней поверхности плодолистиков, а также в нектарных и стигматических сосочках (рис. 6E и 7E, F).
В ранних цветочных почках некоторые мелкие шовные идиобласты наблюдались исключительно в мезофилле органов околоцветника (рис. 5А–С). Швы становились больше и особенно многочисленны по направлению к абаксиальной поверхности чашелистиков и лепестков по мере приближения цветения (рис. 6D, F и 7D). На промежуточных стадиях развития в гинецее и стаминодиях также появлялись маленькие шовные идиобласты, которые становились крупнее к цветению. Было замечено, что несколько рассеянных швов были распределены в базальной области завязи и в локулах на средней высоте, ограниченных боковыми стенками плодолистика (рис.6Г, Д). Более того, присутствие рафидов стало более заметным в верхней части локул и в столбике, в основном в периферических слоях клеток плодолистиков (рис. 6F–H и 7C).
Было замечено, что несколько рассеянных швов были распределены в базальной области завязи и в локулах на средней высоте, ограниченных боковыми стенками плодолистика (рис.6Г, Д). Более того, присутствие рафидов стало более заметным в верхней части локул и в столбике, в основном в периферических слоях клеток плодолистиков (рис. 6F–H и 7C).
ОБСУЖДЕНИЕ
Развитие Acervulus в
Gaussia attenuata Наши наблюдения за ацервулюсом у G. attenuata выявили наличие по крайней мере одного ассоциированного прицветника, структуры, не обнаруженной в предыдущих исследованиях этого рода (Quero, 1982; Quero and Read, 1986; Uhl and Dransfield, 1987; Moya ). и другие., 1991; Дрансфилд и Уль, 1998 г.; Дрансфилд и др. , 2008). Этот прицветник стягивает ацервулюс и, предположительно, может защищать цветочные меристемы на ранних стадиях их дифференциации, как это уже было предложено Ortega and Stauffer (2011) для Hyophorbe lagenicaulis. Существование более одного стягивающего прицветника в ацервулюсе G. attenuata (наблюдаемое для одного ацервулюса в настоящем исследовании) было ранее предложено Uhl и Moore (1978), но потребуются дальнейшие исследования, чтобы лучше понять этот признак в цветочном кластере Chamaedoreeae.
Существование более одного стягивающего прицветника в ацервулюсе G. attenuata (наблюдаемое для одного ацервулюса в настоящем исследовании) было ранее предложено Uhl и Moore (1978), но потребуются дальнейшие исследования, чтобы лучше понять этот признак в цветочном кластере Chamaedoreeae.
Последовательность развития G. attenuata acervulus показывает, что, как и у большинства арекоидных пальм, этот вид является выступающим. Дихогамия была интерпретирована как механизм, снижающий самооплодотворение (Bertin and Newman, 1993) и способствующий ауткроссингу (Henderson, 2002) и, следовательно, гетерозису или гибридной силе (Freeman et al. , 1997). Хотя было показано, что дихогамия развивалась по сложной схеме внутри трибы арекоидов Areceae (в подтрибе Arecinae), с рядом различных протогинных и протандрических линий (Loo et al., 2006), выдающийся характер G. attenuata соответствует преобладающему состоянию в семействе пальмовых. Интересно отметить, что эта тенденция противоположна у однодольных в целом, у которых протогиния более распространена (Bertin, Newman, 1993).
Интересно отметить, что эта тенденция противоположна у однодольных в целом, у которых протогиния более распространена (Bertin, Newman, 1993).
Морфо-анатомические особенности мужских и женских цветков
Хотя функции кремнеземистых телец, шовных и дубильных идиобластов у покрытосеменных до сих пор не ясны, некоторые исследования предполагают их роль в качестве хранилищ метаболических отходов или в предотвращении травоядности (Uhl and Moore, 1973, 1977; Prychid and Rudall, 1999; Дрансфилд и др., 2008). Цветочные ткани G. attenuata часто содержали дубильные вещества и рафиды. Кроме того, мы наблюдали наличие стегм и масляных тел. Стегмы или кремнеземные клетки распространены на ладонях, и ранее наблюдалось, что они связаны с сосудистыми пучками у Chamaedoreeae (Tomlinson, 1961; Tomlinson et al. , 2011), хотя о них никогда не сообщалось в репродуктивных органах. Что касается масляных телец, то эти структуры редко наблюдаются в цветках однодольных растений, и на сегодняшний день о них никогда не сообщалось в этом контексте на пальмах (Lersten et al. , 2006 г.). Необходимо провести дальнейшие исследования, чтобы охарактеризовать точный состав и распределение этих клеточных компонентов в семье. Насколько мы могли заметить, различные типы наблюдаемых клеточных включений появлялись в начале развития, но демонстрировали контрастное распределение во времени и в пространстве. Нефтяные тела явно присутствовали на ранних стадиях развития и швы/стегмы на более поздних стадиях. Этот образец может представлять собой эффективный механизм противодействия травоядности цветов, что подтверждает наблюдения Ула и Мура (1977), которые предположили, что дубильные вещества сдерживают питание и что швы вместе с волокнистыми пучками образуют физический барьер.Также будет интересно исследовать важность клеточных включений у различных членов трибы по отношению к их посетителям-насекомым.
, 2006 г.). Необходимо провести дальнейшие исследования, чтобы охарактеризовать точный состав и распределение этих клеточных компонентов в семье. Насколько мы могли заметить, различные типы наблюдаемых клеточных включений появлялись в начале развития, но демонстрировали контрастное распределение во времени и в пространстве. Нефтяные тела явно присутствовали на ранних стадиях развития и швы/стегмы на более поздних стадиях. Этот образец может представлять собой эффективный механизм противодействия травоядности цветов, что подтверждает наблюдения Ула и Мура (1977), которые предположили, что дубильные вещества сдерживают питание и что швы вместе с волокнистыми пучками образуют физический барьер.Также будет интересно исследовать важность клеточных включений у различных членов трибы по отношению к их посетителям-насекомым.
Септальные нектарники встречаются исключительно у однодольных растений, и о них сообщалось во многих таксонах пальм. Однако мало что известно об их морфологии и анатомии, а исследования структуры и развития были проведены только для нескольких таксонов Arecaceae (Stauffer et al. , 2009). Кроме того, нектарники в пестиках мужских цветков пальмы были зарегистрированы только для нескольких видов, таких как Euterpe precatoria (Küchmeister et al., 1997) и Hyophorbe lagenicaulis (Uhl, 1978). Наличие заметно дифференцированного нектарника как в мужских, так и в женских цветках предполагает склонность к опылению насекомыми. В случае Synechanthus warscewiczianus Зифке и Бернал (2004) на основании наблюдений за производством нектара, внешним видом соцветий и многочисленными посетителями насекомых сделали вывод, что этот вид был энтомофилом. Аналогично, о наличии перегородочного нектарника и базипетального созревания ацервулюса сообщалось для Hyophorbe indica (Uhl, 1978).Вышеупомянутые признаки (развитие базипетального цветка, протандрия, клеточные включения и секреция нектара) предполагают взаимодействие между цветками G. attenuata и его предполагаемыми опылителями.
, 2009). Кроме того, нектарники в пестиках мужских цветков пальмы были зарегистрированы только для нескольких видов, таких как Euterpe precatoria (Küchmeister et al., 1997) и Hyophorbe lagenicaulis (Uhl, 1978). Наличие заметно дифференцированного нектарника как в мужских, так и в женских цветках предполагает склонность к опылению насекомыми. В случае Synechanthus warscewiczianus Зифке и Бернал (2004) на основании наблюдений за производством нектара, внешним видом соцветий и многочисленными посетителями насекомых сделали вывод, что этот вид был энтомофилом. Аналогично, о наличии перегородочного нектарника и базипетального созревания ацервулюса сообщалось для Hyophorbe indica (Uhl, 1978).Вышеупомянутые признаки (развитие базипетального цветка, протандрия, клеточные включения и секреция нектара) предполагают взаимодействие между цветками G. attenuata и его предполагаемыми опылителями.
Половая дифференциация и ее эволюционное значение
Тот факт, что все 107 родов подсемейства пальм Arecoideae являются однодомными, за исключением двух раздельнополых родов, Chamaedorea и Wendlandiella , убедительно свидетельствует о том, что общий предок этой группы также был однодомным.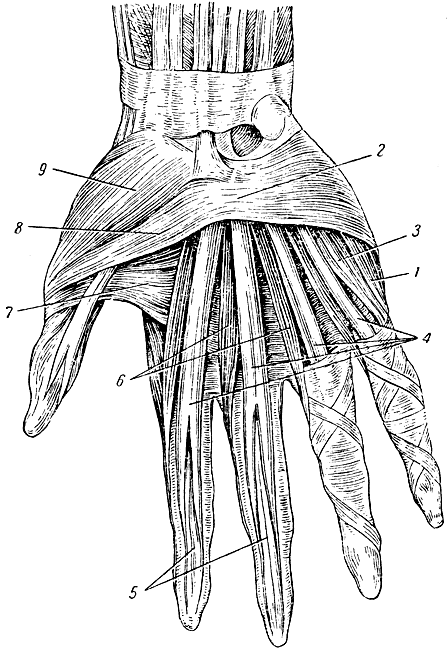 Более того, учитывая, что два вышеупомянутых рода, вероятно, представляют собой независимые эволюционные переходы от однодомного к двудомному в пределах одной и той же трибы, представляется вероятным, что Chamaedoreeae более предрасположены, чем другие арекоидные пальмы, к эволюции однополых растений (Cuenca et al. , 2009). . Важно отметить, что молекулярные механизмы, управляющие однополостью цветка, почти наверняка произошли от общего однодомного предка пяти существующих родов. Таким образом, даже несмотря на то, что в раздельнополых линиях произойдут геномные перестройки, создающие генетическую систему с отдельными мужскими и женскими растениями, вполне вероятно, что в этих кладах будут действовать те же гены детерминации пола, что и в однодомных.На морфо-анатомическом уровне сравнение Gaussia , Hyophorbe и Chamaedorea (т.е. трех родов трибы, по которым в настоящее время имеются данные) обнаруживает в целом сильное сходство в паттернах половой дифференциации.
Более того, учитывая, что два вышеупомянутых рода, вероятно, представляют собой независимые эволюционные переходы от однодомного к двудомному в пределах одной и той же трибы, представляется вероятным, что Chamaedoreeae более предрасположены, чем другие арекоидные пальмы, к эволюции однополых растений (Cuenca et al. , 2009). . Важно отметить, что молекулярные механизмы, управляющие однополостью цветка, почти наверняка произошли от общего однодомного предка пяти существующих родов. Таким образом, даже несмотря на то, что в раздельнополых линиях произойдут геномные перестройки, создающие генетическую систему с отдельными мужскими и женскими растениями, вполне вероятно, что в этих кладах будут действовать те же гены детерминации пола, что и в однодомных.На морфо-анатомическом уровне сравнение Gaussia , Hyophorbe и Chamaedorea (т.е. трех родов трибы, по которым в настоящее время имеются данные) обнаруживает в целом сильное сходство в паттернах половой дифференциации. Возможно, наиболее заметное расхождение, которое можно увидеть между родами, заключается в степени дифференциации стаминодий: исследования, проведенные Askgaard et al. (2008) показал, что у некоторых (но не у всех) видов Chamaedorea женских цветков отсутствуют какие-либо следы стерильного андроцея.Эта ситуация контрастирует с таковой у Hyophorbe и Gaussia , у которых стаминодии всегда четко различимы, несмотря на относительно раннюю задержку развития. Различия между видами Chamaedorea по наличию или отсутствию стаминодий могут отражать генетические изменения, произошедшие с момента появления линии Chamaedorea . Тот факт, что половой диморфизм неоднократно развивался у покрытосеменных и несколько раз у пальм, означает, что разные генетические детерминанты, вероятно, будут применяться в каждой линии (Diggle et al., 2011). На сегодняшний день единственными двумя видами покрытосеменных, для которых была выполнена функциональная идентификация генов детерминации пола, является кукуруза (DeLong et al.
Возможно, наиболее заметное расхождение, которое можно увидеть между родами, заключается в степени дифференциации стаминодий: исследования, проведенные Askgaard et al. (2008) показал, что у некоторых (но не у всех) видов Chamaedorea женских цветков отсутствуют какие-либо следы стерильного андроцея.Эта ситуация контрастирует с таковой у Hyophorbe и Gaussia , у которых стаминодии всегда четко различимы, несмотря на относительно раннюю задержку развития. Различия между видами Chamaedorea по наличию или отсутствию стаминодий могут отражать генетические изменения, произошедшие с момента появления линии Chamaedorea . Тот факт, что половой диморфизм неоднократно развивался у покрытосеменных и несколько раз у пальм, означает, что разные генетические детерминанты, вероятно, будут применяться в каждой линии (Diggle et al., 2011). На сегодняшний день единственными двумя видами покрытосеменных, для которых была выполнена функциональная идентификация генов детерминации пола, является кукуруза (DeLong et al. , 1993; Chuck et al. , 2007; Acosta et al. , 2009). и дыни (Boualem и др. , 2008; Martin и др. , 2009). В случае Arecaceae были получены доказательства сохранения по крайней мере некоторых функций генов, связанных с ABC-моделью цветков (Coen and Meyerowitz, 1991; Pelaz et al., 2000) у африканской масличной пальмы Elaeis guineensis (Adam et al. , 2007). Однако до сих пор не установлено прямой связи между генами, определяющими гомеозисную идентичность цветочных мутовок, и процессом детерминации пола per se . В случае Chamaedoreeae дальнейшие исследования, направленные на характеристику клеточных изменений, связанных с остановкой стерильности органов, должны помочь пролить свет на процесс половой дифференцировки. Подход такого типа уже использовался с финиковой пальмой ( Phoenix dactylifera ), чтобы продемонстрировать, что деление клеток прекращается в стерильном андроцее и гинецее женских и мужских цветков соответственно при отсутствии какой-либо потери ядерной целостности (Daher et al.
, 1993; Chuck et al. , 2007; Acosta et al. , 2009). и дыни (Boualem и др. , 2008; Martin и др. , 2009). В случае Arecaceae были получены доказательства сохранения по крайней мере некоторых функций генов, связанных с ABC-моделью цветков (Coen and Meyerowitz, 1991; Pelaz et al., 2000) у африканской масличной пальмы Elaeis guineensis (Adam et al. , 2007). Однако до сих пор не установлено прямой связи между генами, определяющими гомеозисную идентичность цветочных мутовок, и процессом детерминации пола per se . В случае Chamaedoreeae дальнейшие исследования, направленные на характеристику клеточных изменений, связанных с остановкой стерильности органов, должны помочь пролить свет на процесс половой дифференцировки. Подход такого типа уже использовался с финиковой пальмой ( Phoenix dactylifera ), чтобы продемонстрировать, что деление клеток прекращается в стерильном андроцее и гинецее женских и мужских цветков соответственно при отсутствии какой-либо потери ядерной целостности (Daher et al. ., 2010). У G. attenuata задержка развития, по-видимому, имеет место на относительно ранней стадии после инициации в случае стаминодий и на гораздо более поздней стадии у пестичных; однако в последнем случае развитие семязачатка отсутствует и заменяется появлением вегетативных тканей, окружающих нектарник.
., 2010). У G. attenuata задержка развития, по-видимому, имеет место на относительно ранней стадии после инициации в случае стаминодий и на гораздо более поздней стадии у пестичных; однако в последнем случае развитие семязачатка отсутствует и заменяется появлением вегетативных тканей, окружающих нектарник.
В заключение, наши данные предоставляют новую полезную информацию о развитии цветков у ранее неизученного рода Chamaedoreeae, которая поможет улучшить наше понимание эволюционных изменений, сопровождавших смену половой системы в этой трибе.Дальнейшие исследования с использованием дополнительных молекулярных гистологических подходов должны предоставить информацию о том, как достигается арест стерильных репродуктивных органов, а развитие геномных ресурсов впоследствии поможет получить представление о лежащих в основе молекулярных событиях.
БЛАГОДАРНОСТИ
Мы очень благодарны Ботаническому центру Монтгомери (США) за доступ к их коллекции живых пальм для этого исследования. Особая благодарность д-ру Патрику Гриффиту, директору Центра, и д-ру Ларри Ноблику, предоставившем фотографию для этой статьи.Мы также признательны доктору Андре Пью (Музей естественной истории, Женева, Швейцария), Шанталь Казевьей и Сесиль Санчес (CRIC, Монпелье, Франция) за их техническую помощь в работе с РЭМ. Д-р Глория Галеано (Instituto de Ciencias Naturales, Universidad Nacional de Colombia) выражает благодарность за постоянную поддержку проекта. Коллектив библиотеки Ботанической консерватории в Женеве выражает благодарность за постоянный доступ к своим коллекциям. Эта работа стала возможной благодаря гранту PhD от IRD Фелипе Кастаньо и основному финансированию со стороны наших учреждений.Мы благодарим Билла Бейкера и анонимного рецензента за полезные комментарии, позволившие нам улучшить рукопись.
Особая благодарность д-ру Патрику Гриффиту, директору Центра, и д-ру Ларри Ноблику, предоставившем фотографию для этой статьи.Мы также признательны доктору Андре Пью (Музей естественной истории, Женева, Швейцария), Шанталь Казевьей и Сесиль Санчес (CRIC, Монпелье, Франция) за их техническую помощь в работе с РЭМ. Д-р Глория Галеано (Instituto de Ciencias Naturales, Universidad Nacional de Colombia) выражает благодарность за постоянную поддержку проекта. Коллектив библиотеки Ботанической консерватории в Женеве выражает благодарность за постоянный доступ к своим коллекциям. Эта работа стала возможной благодаря гранту PhD от IRD Фелипе Кастаньо и основному финансированию со стороны наших учреждений.Мы благодарим Билла Бейкера и анонимного рецензента за полезные комментарии, позволившие нам улучшить рукопись.
ЦИТИРОВАННАЯ ЛИТЕРАТУРА
, , , и другие. tasselseed1 представляет собой липоксигеназу, влияющую на передачу сигналов жасмоновой кислоты при определении пола кукурузы.
323
(стр.262
—265
), , , , , .Сложность репродуктивного развития африканской масличной пальмы ( Elaeis guineensis , Arecaceae)
,American Journal of Botany
,2005
, vol.92
(стр.1836
—1852
), , , , , , .Функциональная характеристика генов коробки MADS, участвующих в определении структуры цветка масличной пальмы
58
(стр.1245
—1259
), , , .Цветочная структура неотропического рода пальм Chamaedorea (Arecoideae, Arecaceae)
,Anales del Jardín Botánico de Madrid
,2008
, vol.65
(стр.197
—210
), , , , , .Новая классификация подсемейства семейства пальмовых (Arecaceae): данные филогении пластидной ДНК
151
(стр.15
—38
), , , и др.Полный филогенетический анализ пальм (Arecaceae) на общем уровне со сравнением подходов супердерева и суперматрицы
58
(стр.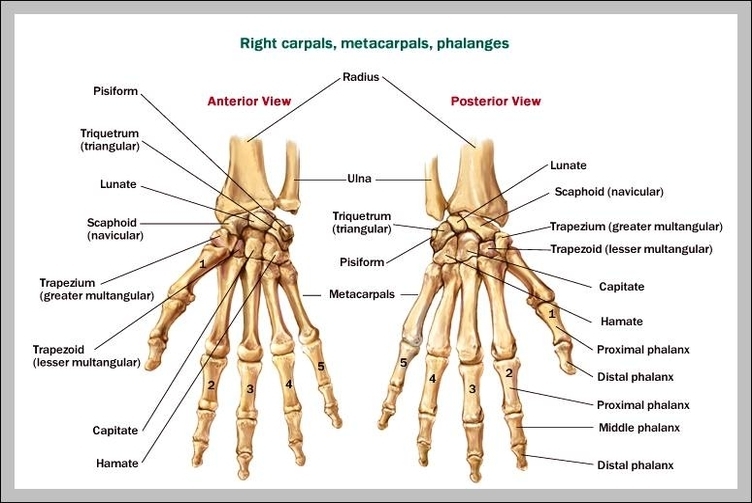
240
—56
), , , и др.Филогенетические взаимоотношения между арекоидными пальмами (Arecaceae: Arecoideae)
,Летопись ботаники
,2011
, том.108
(стр.1417
—1432
).Эволюция репродуктивных систем растений: как часто переходы необратимы?
,Proceedings of the Royal Society B: Biological Sciences
,2013
, vol.280
стр.20130913
, .Дихогамия покрытосеменных
,Ботанический обзор
,1993
, том.59
(стр.112
—152
), , , и др.Законсервированная мутация в ферменте биосинтеза этилена приводит к андромоноэции в дынях
321
(стр.836
—838
), , , , .МикроРНК tasselseed4 кукурузы контролирует детерминацию пола и судьбу клеток меристемы путем нацеливания на Tasselseed6/индетерминантный колосок1
39
(стр.1517
—1521
), .Война мутовок: генетические взаимодействия, контролирующие развитие цветка
,Природа
,1991
, том.
353
(стр.31
—37
), .Филогения пальмовой трибы Chamaedoreeae (Arecaceae) на основе последовательностей пластидной ДНК
,Систематическая ботаника
,2007
, vol.32
(стр.250
—263
), , .Датированная филогения пальмового племени Chamaedoreeae подтверждает эоценовое расселение между Африкой, Северной и Южной Америкой
46
(стр.760
—775
), , .Филогения и эволюция морфологических признаков трибы Chamaedoreeae (Arecaceae)
,Taxon
,2009
, vol.58
(стр.1092
—1108
), , , и др.Остановка клеточного цикла характеризует переход от двуполой цветочной почки к однополому цветку у Phoenix dactylifera
,Annals of Botany
,2010
, vol.106
(стр.255
—266
), , .Ген определения пола TASSELSEED2 кукурузы кодирует короткоцепочечную алкогольдегидрогеназу, необходимую для аборта цветочных органов на определенной стадии
74
(стр.
757
—768
), , , и др.Множественные процессы развития лежат в основе половой дифференциации покрытосеменных растений
,Тенденции генетики
,2011
, vol.27
(стр.368
—376
), ..Palmae
,Семейства и роды сосудистых растений. Цветущие растения. Однодольные
,1998
Berlin
Springer
(стр.306
—389
), , , , , .Новая филогенетическая классификация семейства пальмовых, Arecaceae
,Kew Bulletin
,2005
, vol.60
(стр.559
—569
), , , , , . ,Роды пальм: эволюция и классификация пальм
,2008
Кью, Великобритания
Kew Publishing
, .Реконструкция предкового цветка покрытосеменных и его первоначальных специализаций
96
(стр.22
—66
), , , , .Половая специализация и предотвращение инбридинга в эволюции раздельнополых
,Ботанический обзор
,1997
, том.
63
(стр.65
—92
), . ,Контрольный список пальм
,2005
Лондон
Королевские ботанические сады, Кью
.Филогенетический анализ арекоидной линии пальм на основе данных о последовательностях пластидной ДНК
23
(стр.189
—204
). ,Эволюция и экология пальм
,2002
Нью-Йорк
New York Botanical Garden Press
, , . ,Полевой справочник по пальмам Америки
,1995
Принстон, Нью-Джерси
Princeton University Press
, , .Цветение, опыление, нектароплоды и нектарники Euterpe precatoria (Arecaceae), пальмы тропических лесов Амазонки
206
(стр.71
—97
), , , , .Масляные тельца в клетках мезофилла листа покрытосеменных растений: обзор и избранный обзор
93
(стр.1731
—1739
), , , .Низкокопийная ядерная ДНК, филогения и эволюция дихогамии у ореховых пальм бетеля и их родственников (Arecinae; Arecaceae)
39
(стр.
598
—618
), , , и др.Индуцированное транспозоном эпигенетическое изменение приводит к определению пола у дыни
,Nature
,2009
, vol.461
(стр.1135
—1138
).Основные группы пальм и их распространение
,Gentes Herbarum
,1973
, vol.11
(стр.27
—141
), , , , .Gaussia Spirituana Moya et Leiva, sp. ноябрь: Уна-Нуэва-Пальма-де-Куба, центральная
,Revista del Botánico Nacional
,1991
, vol.12
(стр.15
—20
), .Онтогенез и строение асервулатного парциального соцветия у Hyophorbe lagenicaulis (Arecaceae; Arecoideae)
,Annals of Botany
,2011
, vol.108
(стр.1517
—1527
), , , , .Для функций идентичности цветочных органов B и C требуются гены SEPALLATA MADS-box
,Nature
,2000
, vol.405
(стр.
200
—203
), .Кристаллы оксалата кальция в однодольных: обзор их строения и систематики
,Анналы ботаники
,1999
, том.84
(стр.725
—739
).Opsiandra gomez-pompae , новый вид из Оахаки, Мексика
,Principes
,1982
, vol.26
(стр.144
—149
), .Редакция рода пальм Gaussia
,Систематическая ботаника
,1986
, vol.11
(стр.145
—154
).Материалы для словаря ботанических терминов III. Соцветия
,Бюллетень Ботанического клуба Торри
,1955
, том.82
(стр.419
—445
), .Половой диморфизм в приобретении и использовании ресурсов: важны как размер, так и время
107
(стр.119
—126
), .Биология флоры и насекомые-посетители подлеска пальмы Synechanthus warscewiczianus на тихоокеанском побережье Колумбии
,Пальмы
,2004
, vol.
48
(стр.33
—41
), , .Цветочная структура в Licuala peltata (Arecaceae: Coryphoideae) с особой ссылкой на архитектуру необычного лабиринтного нектарника
,Ботанический журнал Линнеевского общества
,2009
, vol.161
(стр.66
—77
), , , и др.Молекулярная филогения пальм рода Chamaedorea, основанная на низкокопийных ядерных генах PRK и RPB2
38
(стр.398
—415
). .Palmae
,Анатомия однодольных
,1961
Oxford
Oxford University Press
, , . ,Анатомия пальм: Arecaceae – Palmae
,2011
Нью-Йорк
Oxford University Press
.Исследования развития Ptychosperma (Palmae). I. Соцветие и соцветие
,American Journal of Botany
,1976
, vol.63
(стр.82
—96
).Цветочная анатомия пяти видов Hyophorbe
,Gentes Herbarum
,1978
, vol.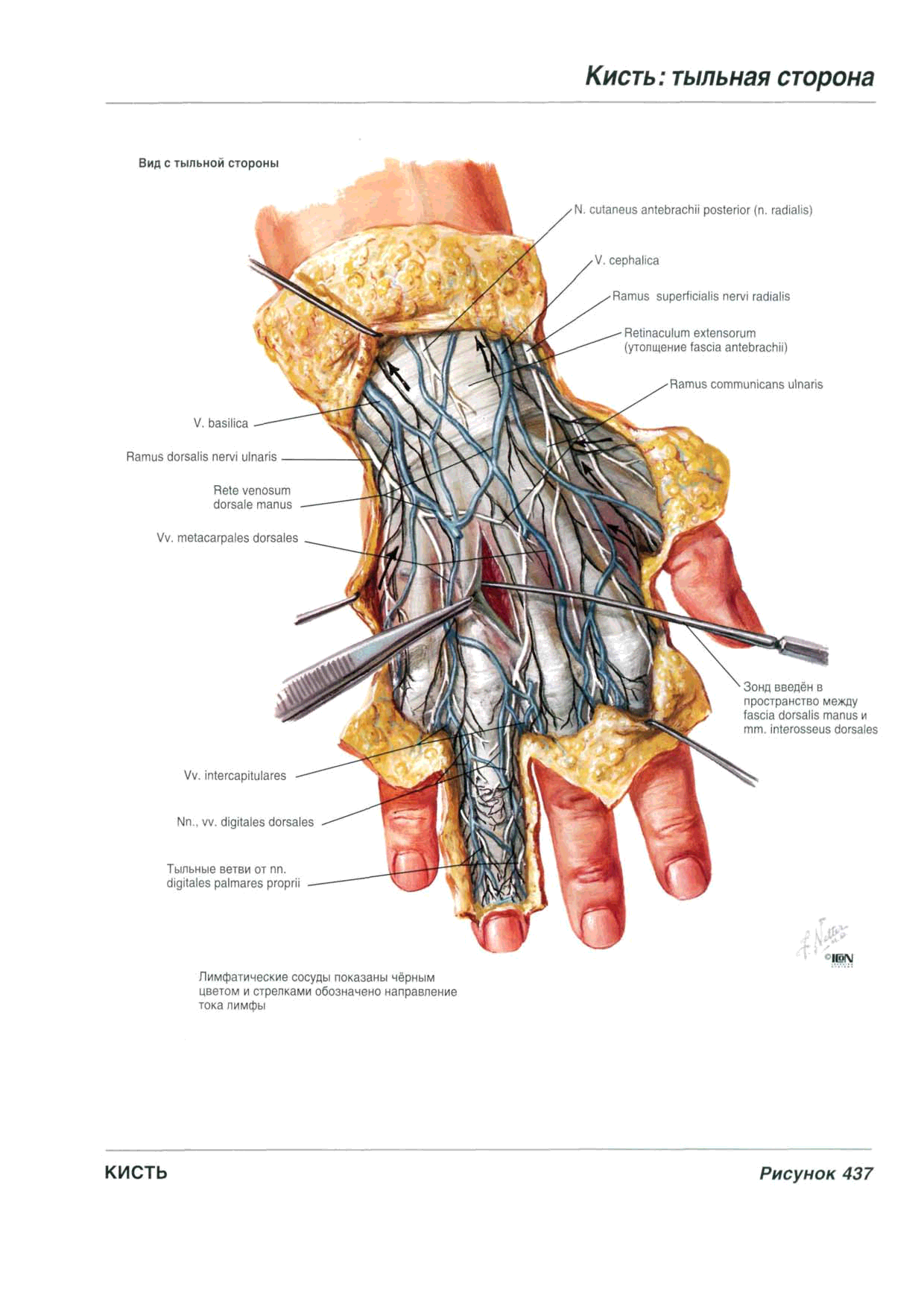
11
(стр.245
—267
), .Гинецей пальмы
,American Journal of Botany
,1971
, vol.58
(стр.945
—992
), . , .Развитие соцветия, андроцея и гинецея применительно к пальмам
,Современные проблемы анатомии растений
,1984
Нью-Йорк
Academic Press
(стр.390
— 0 ,
Genera palmarum
,1987
Лоуренс, Канзас
Allen Press
, .Защита пыльцы и семяпочек на ладонях
,Principes
,1973
, vol.17
(стр.111
—149
), .Взаимосвязь соцветия, структуры цветка и анатомии цветка с опылением на некоторых пальмах
9
(стр.170
—190
), .Строение ацервулюса, соцветия хамедоновидной пальмы
,American Journal of Botany
,1978
, vol.65
(стр.197
—204
), , , , , ., , , .Филогенетические отношения между ладонями: Кладистический анализ морфологического и хлоропластового ДНК Ограничение площадки
,Монокотилоны, систематики и эволюция
,1995
KEW
Королевские ботанические сады
(стр.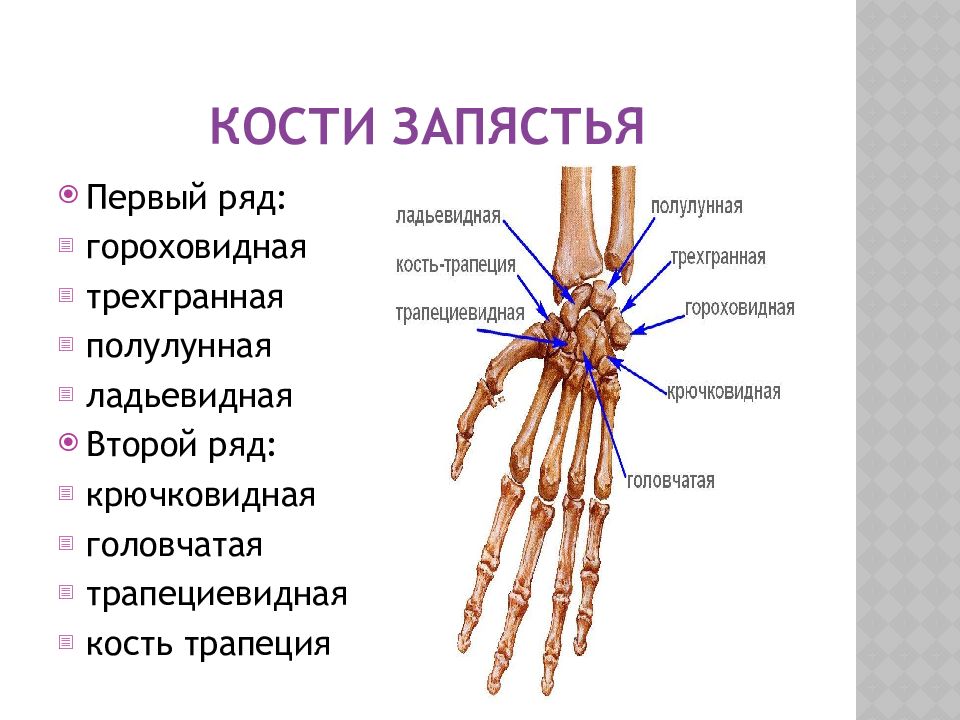
623
—661
), .Филогенетический анализ двудомности однодольных
,American Naturalist
,2000
, vol.155
(стр.46
—58
)© Автор 2014.Опубликовано Oxford University Press от имени Annals of Botany Company. Все права защищены. Для получения разрешений отправьте электронное письмо по адресу: [email protected]
.Строение масличной пальмы
Широко известная как масличная пальма, Elaeis guineensis и многие ее составные части использовались людьми по крайней мере с 3000 г. до н.э. Родом из Западной и Центральной Африки, около 44 стран в настоящее время используют это растение для производства пальмового масла.
Пальмовое масло (Изображение: Лени Кауфман/China Dialogue) Пальмовое масло является наиболее широко используемым растительным маслом в мире: в 2019 году во всем мире было произведено 76 миллионов тонн.Несмотря на это, масло составляет лишь 10% биомассы масличной пальмы.
Итак, что мы знаем о других частях этой тропической культуры, которая на самом деле не дерево, а большая древесная трава?
Корни
Экономическая продолжительность жизни масличной пальмы составляет около 25-30 лет, после чего урожай снижается, и пальма становится слишком высокой для эффективного сбора урожая. По прошествии своего расцвета зрелые пальмы срубают, чтобы освободить место для новых.
Выкорчевывание пальм часто связано с использованием тяжелой техники или незаконными методами рубки и сжигания, которые серьезно разрушают почву, делая ее уязвимой для эрозии и менее способной поглощать воду.Это увеличивает риск крупных наводнений в близлежащих районах в периоды сильных дождей.
Наличие тяжелой техники также снижает способность почвы сохранять воду и кислород, что приводит к анаэробным условиям, которые не могут поддерживать питательные вещества и органические вещества, способствующие разложению и сохраняющие почву здоровой.
Корни масличной пальмы (Изображение: Лени Кауфман/China Dialogue) Хотя в настоящее время это противоречит передовой отраслевой практике, многие новые плантации исторически создавались на торфяниках, которые были расчищены и осушены, что привело к разрушению богатых экосистем и жизненно важных запасов углерода.
Воздействие деятельности плантаций простирается от подземных до близлежащих источников воды. Агрохимикаты, такие как удобрения, пестициды и гербициды, используются на плантациях пальмового масла. Химические стоки в близлежащие ручьи, реки и озера могут стимулировать производство водорослей, которые быстро растут и лишают других водных организмов кислорода в процессе, называемом эвтрофикацией.
Плантации масличной пальмы невероятно водоемки: в среднем на одну масличную пальму в Малайзии уходит около 100 литров воды каждый день.Около 243 000 литров — или десятая часть олимпийского бассейна — требуется для производства тонны свежих фруктовых связок.
Плодовые гроздья
В возрасте трех лет масличные пальмы начинают приносить около дюжины гигантских плодовых кистей, каждая из которых может весить до 30 кг. Из корешков этих ярких гроздей вырастает до 3000 отдельных плодов, дающих масло. Масличные пальмы обеспечивают постоянный урожай, так как каждая отдельная пальма дает урожай свежих фруктов каждые две недели.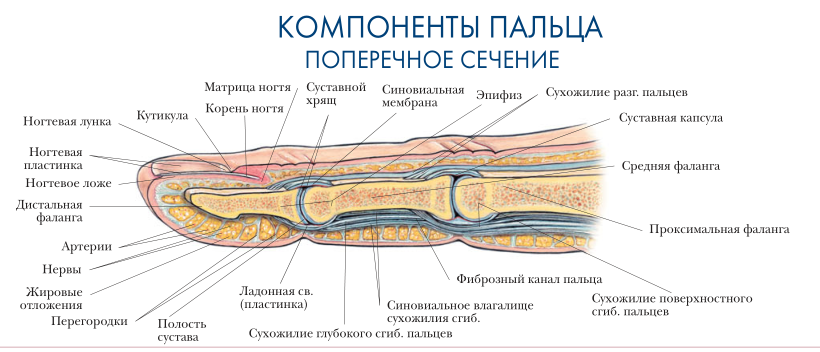
Путь независимых мелких фермеров часто заканчивается гроздью свежих фруктов, которые они продают на местные мельницы для переработки.
На заводе по производству пальмового масла собранные грозди обмолачивают, отделяя плоды от гроздей. Остается скелет фруктовой грозди, который можно переработать в мульчу или использовать для производства биотоплива. Однако современные методы часто являются дорогостоящими, что затрудняет обработку для мелких землевладельцев.
Листья
Исторически сложилось так, что непослушные листья масличной пальмы использовались людьми для плетения кустарных корзин, изготовления крыш для домов или лечения ран с помощью традиционной медицины.Но массовое производство пальмового масла изменило ситуацию, и листья ежедневно выбрасываются как отходы.
Вайя масличной пальмы (Изображение: Лени Кауфман/China Dialogue) Вайя масличной пальмы имеет длину в среднем три метра, при этом ежегодно обрезается около 24 ветвей на каждом дереве, накапливая огромное количество биомассы.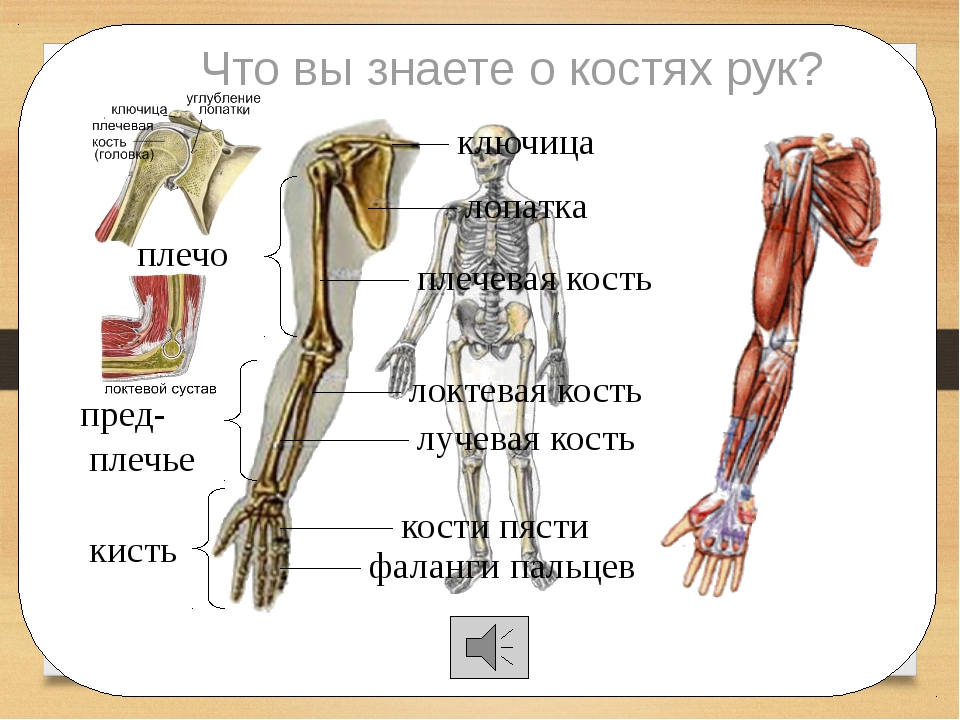
Изобретательные способы использования избыточных листьев масличной пальмы помимо мульчи и биотоплива являются обычным явлением на плантациях. Благодаря тщательной укладке и штабелированию листьев среди рядов масличных пальм они начинают гнить, образуя щит для удержания влаги в почве, а также уменьшая атаки вредителей и пополняя почву питательными веществами.
На многих плантациях эти стеки листьев являются единственной растительностью на земле и могут обеспечить столь необходимую среду обитания для животных, живущих в ландшафте с масличными пальмами. Кобры, вараны, беспозвоночные и грызуны обычно встречаются в этих ветвях.
Плоды Плоды масличной пальмы (Изображение: Лени Кауфман/Китайский диалог)
Африканский пальмовый долгоносик ( Rhynchophorus phoenicis ) является насекомым-опылителем масличной пальмы и жизненно важен для получения плодотворных урожаев.До того, как оно было завезено в Юго-Восточную Азию, урожайность масличной пальмы была на 40% ниже, чем сегодня.
Через шесть месяцев после опыления плоды масличной пальмы достигают размера сливы и меняют цвет от темно-фиолетового до огненно-оранжевого и красного.
Каждый плод содержит около 50% масла, и как мякоть (известная как мезокарпий), так и ядро могут выделять масло.
Ядро, расположенное в центре плода, является источником пальмоядрового масла. Именно это темно-желтое масло обычно используется для производства моющих средств, косметики, пластмасс и химикатов.Сырое пальмовое масло, извлеченное из мезокарпия плода, дает масло темно-красного цвета, которое содержится в различных продуктах.
Ствол
Традиционно со стволов удаляли оставшиеся основания листьев, так как считалось, что они содержат паразитические растения или болезни. Совсем недавно производители начали покидать эти базы, чтобы поддержать почвообразование и биоразнообразие на плантациях масличной пальмы. Растения могут расти во взвешенных почвах, которые собираются в полостях между листьями и стеблем, где также могут найти пристанище сообщества беспозвоночных.
Помимо того, что ствол масличной пальмы может служить хозяином для биоразнообразия, сердцевина ствола масличной пальмы имеет мясистую текстуру и часто поедается слонами, орангутангами и дикими кабанами, которые бродят по молодым плантациям в поисках еды.
Ствол масличной пальмы (Изображение: Лени Кауфман/China Dialogue)Стволы масличной пальмы имеют почти идеально цилиндрическую форму, что делает их удобными для распиливания и очистки. Однако, согласно исследованиям, только одна треть радиуса ствола — внешний слой — достаточно прочна, чтобы ее можно было использовать в качестве цельной древесины.Использование древесины масличной пальмы было минимальным и может быть дорогостоящим, поскольку ее низкое качество требует серьезной обработки, чтобы сделать ее пригодной для изготовления мебели или строительства.
Имея около 17 миллионов гектаров зрелых пальмовых плантаций на экваторе, потенциальное использование древесины масличной пальмы является ключевой областью исследований и разработок, поскольку отрасль пытается сделать цикл производства пальмового масла максимально устойчивым. Post Categories: Разное
Post Categories: Разное
 By alexxlab
By alexxlab
