
Стили (формы) бонсай — Империя Бонсай
Стили (формы) бонсай
Стили, лежащие в основе классификации бонсай, напоминают различные формы деревьев в природе. Эти стили могут трансформироваться в процессе индивидуального творческого осмысления, т.е. деревья не должны обязательно соответствовать какой-либо конкретной форме.
Тем не менее, важность этих стилей состоит в том, что они помогают получить общее представление о формах деревьев и служат в качестве ориентира для успешного формирования бонсай.
Веерный стиль или метла (хокидати)
Веерный стиль подходит для лиственных деревьев с длинным и тонким ветвлением. Ствол — прямой и вертикальный, однако он не доходит до верхушки дерева, так как разветвляется в разные стороны в точке, находящейся на расстоянии около 1/3 от общей высоты дерева. Ветви и листья образуют густую шарообразную крону, которая и в зимнее время поражает воображение.
Формальный вертикальный стиль (тёккан)
Формальный вертикальный стиль весьма распространен в бонсай. Такая форма дерева часто встречается в природе, особенно если дерево растет в солнечном месте, не конкурируя с другими деревьями. Ствол дерева в этом стиле должен обладать хорошей сбежистостью, т.е. постепенно сужаться от основания к макушке. Ветки должны появляться на расстоянии около 1/4 от общей высоты ствола.
Такая форма дерева часто встречается в природе, особенно если дерево растет в солнечном месте, не конкурируя с другими деревьями. Ствол дерева в этом стиле должен обладать хорошей сбежистостью, т.е. постепенно сужаться от основания к макушке. Ветки должны появляться на расстоянии около 1/4 от общей высоты ствола.
Неформальный вертикальный стиль (мойоги)
Неформальный вертикальный стиль распространен как в природе, так и в искусстве бонсай. Ствол растет в целом вертикально, но имеет форму, напоминающую английскую букву «S», а на внешней стороне каждого изгиба растут ветки. Сбежистость ствола должны быть хорошо видна, т.е. основание ствола должно быть толще, чем его верхняя часть.
Наклонный стиль бонсай (сяккан)
Если ветра дуют преимущественно в одном направлении, или дерево растет в тени, оно вынуждено сгибаться в сторону солнца и наклоняться в определенном направлении. В бонсай дерево в наклонном стиле должно расти под углом около 60 — 80 градусов по отношению к поверхности грунта. Со стороны, противоположной наклону, корни должны быть мощные, чтобы удерживать дерево. На другой стороне корни не так хорошо развиты. Первая ветвь, как правило, растет на стороне, противоположной наклону, чтобы обеспечить визуальную сбалансированность композиции. Ствол может быть слегка изогнутыми или совершенно прямым, но при этом более толстым у основания, чем в верхней части дерева.
Со стороны, противоположной наклону, корни должны быть мощные, чтобы удерживать дерево. На другой стороне корни не так хорошо развиты. Первая ветвь, как правило, растет на стороне, противоположной наклону, чтобы обеспечить визуальную сбалансированность композиции. Ствол может быть слегка изогнутыми или совершенно прямым, но при этом более толстым у основания, чем в верхней части дерева.
Каскадный стиль (кенгай)
Дерево, растущее в природе на крутом обрыве, может согнуться под воздействием тяжести снега, упавших камней или других факторов. В бонсай сохранять такое направление роста дерева может быть непросто, поскольку это противоречит естественному стремлению дерева расти вертикально вверх. Каскадный бонсай сажают в высокие горшки. Основание дерева может быть относительно вертикальным, но потом ствол загибается вниз. Макушка дерева, как правило, находится выше края горшка, а остальные ветки чередуются справа и слева на внешних изгибах зигзагообразного каскадного ствола. Ветки расположены горизонтально, чтобы обеспечивать визуальную сбалансированность композиции.
Ветки расположены горизонтально, чтобы обеспечивать визуальную сбалансированность композиции.
Полукаскадный стиль (хан кенгай)
Полукаскадный стиль, как и каскадный, встречается в природе на скалах берегах водёмов. Нижняя часть ствола растет вертикально, а затем наклоняется вниз и в сторону. В отличие от каскадного стиля, в этом случае каскад оканчивается не ниже дна горшка. Макушка самого дерева, как правило, возвышается над верхним краем горшка.
Литературный (богемный) стиль (бунзинги)
В природе такой стиль дерева можно найти в местах, где деревья растут так густо, а конкуренция между ними настолько ожесточенная, что дерево выживет, только если вырастет выше других. Ствол — вертикальный, но несколько кривой, и полностью без веток, так как только верхушка дерева освещается солнцем. Для визуального состаривания дерева некоторые ветви «заджиниваются» (искусственно умертвляются). Если кора удаляется с одной стороны ствола – это называется «сяри». Эти приемы призваны продемонстрировать борьбу дерева за выживание. Бонсай в этом стиле часто сажают в маленькие круглые горшки.
Эти приемы призваны продемонстрировать борьбу дерева за выживание. Бонсай в этом стиле часто сажают в маленькие круглые горшки.
Стиль дерево, согнутое ветром (фукинагаси)
Этот стиль также ярко демонстрирует деревья, которые борются за выживание. Ветви, а также ствол растут по направлению доминирующих ветров. Ветви могут расти по всей окружности ствола, но в конечном итоге сгибаются в одну сторону.
Стиль двойной ствол (сёкан)
Двойной ствол распространен в природе, но на самом деле не так часто встречается в искусстве бонсай. Обычно оба ствола растут из одной корневой системы, но возможен вариант, когда маленький ствол растет из большого ствола чуть выше основания. Два ствола отличаются толщиной и длиной, более толстый и взрослый растет почти вертикально, а меньший — немного наклонно. При этом они образуют единую крону.
Многоствольный стиль (кабудати)
По сути многоствольный стиль аналогичен двухствольному, но с 3 или более стволами. Все они растут из одной корневой системы. Стволы образуют единую крону, при этом самый толстый из них является самым высоким.
Все они растут из одной корневой системы. Стволы образуют единую крону, при этом самый толстый из них является самым высоким.
Стиль роща или групповая посадка (йосе-уэ)
Стиль роща похож на многоствольный, но разница в том, что роща состоит из нескольких отдельных деревьев. Наиболее развитые деревья высаживают в середине большого мелкого поддона. По бокам сажается несколько небольших деревьев, которые все вместе образуют единую крону. Деревья высаживают не по прямой, а в произвольном порядке, чтобы роща выглядела реалистично и естественно.
Стиль корни на скале (секийою)
В каменистой местности деревья вынуждены искать своими корнями богатую питательными веществами почву, которая иногда скапливается в трещинах и углублениях. Пока корни не попали в землю, они защищают себя от солнца, покрываясь грубой корой. В случае бонсай, корни растут, оплетая камень, а затем погружаются в грунт в горшке, поэтому уход за таким деревом на самом деле не отличается от ухода за деревом в любом другом стиле бонсай.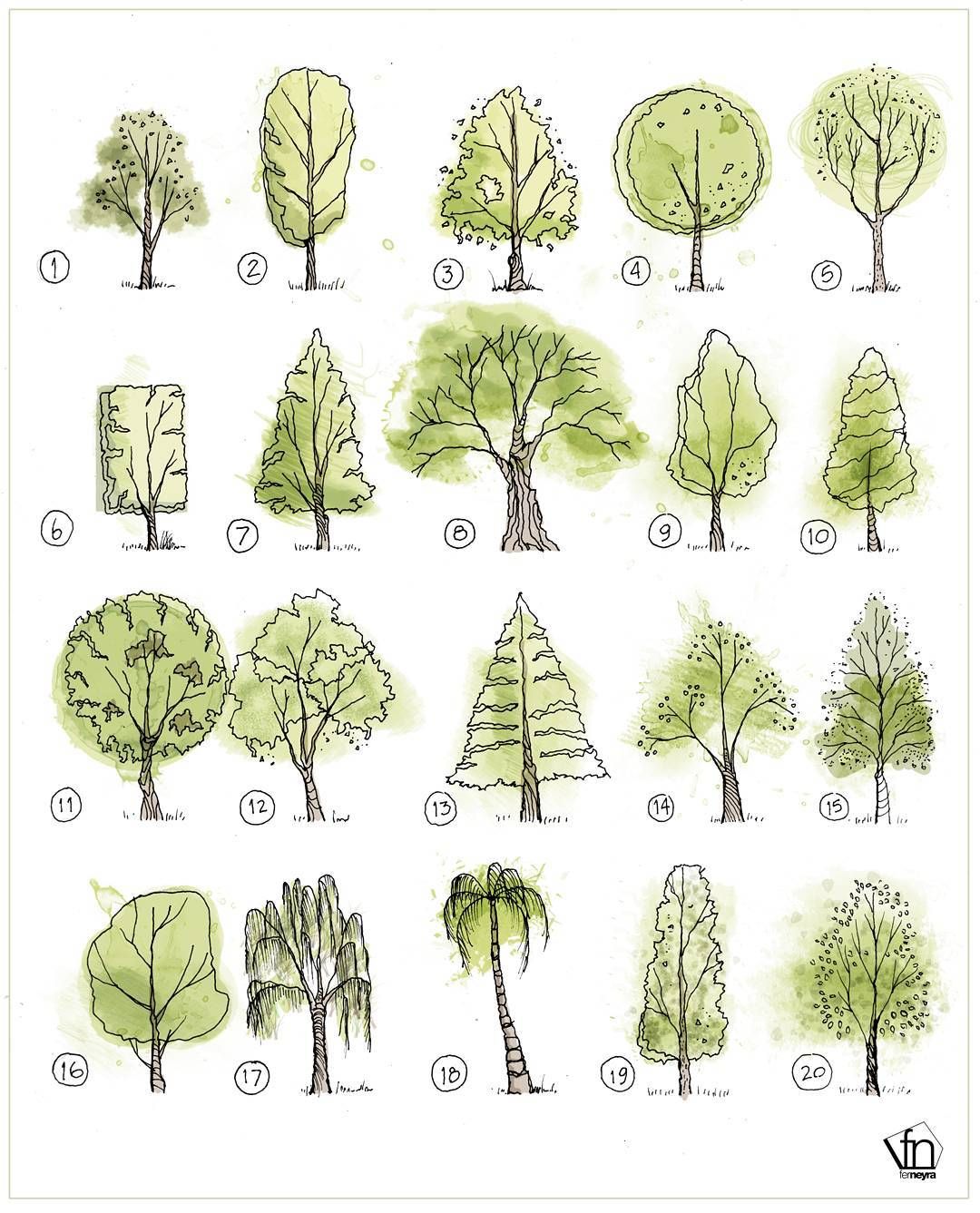 Для этого стиля подходят, в частности, можжевельники и фикусы.
Для этого стиля подходят, в частности, можжевельники и фикусы.
Дерево на камне (исицуки)
У деревьев, сформированных в этом стиле, корни растут в трещинах и полостях камня. Это означает, что у них ограниченные возможности для роста и получения питательных веществ. Деревья, растущие в камне, никогда не выглядят действительно здоровыми, что дополнительно демонстрирует сложности их борьбы за выживание. Их необходимо часто поливать и удобрять, поскольку у них мало места для хранения влаги и питательных веществ. Камень, в котором растет такой бонсай, часто помещают в мелкий поддон, который иногда заполняется водой или засыпается мелким гравием.
Стиль упавшее дерево (икадабуки)
Иногда упавшее дерево может выжить, а его ветки начинают расти вверх. Старая корневая система может обеспечивать эти ветви достаточными для выживания питательными веществами. Через некоторое время у них начинают расти новые корни, в конце концов замещающие старую корневую систему.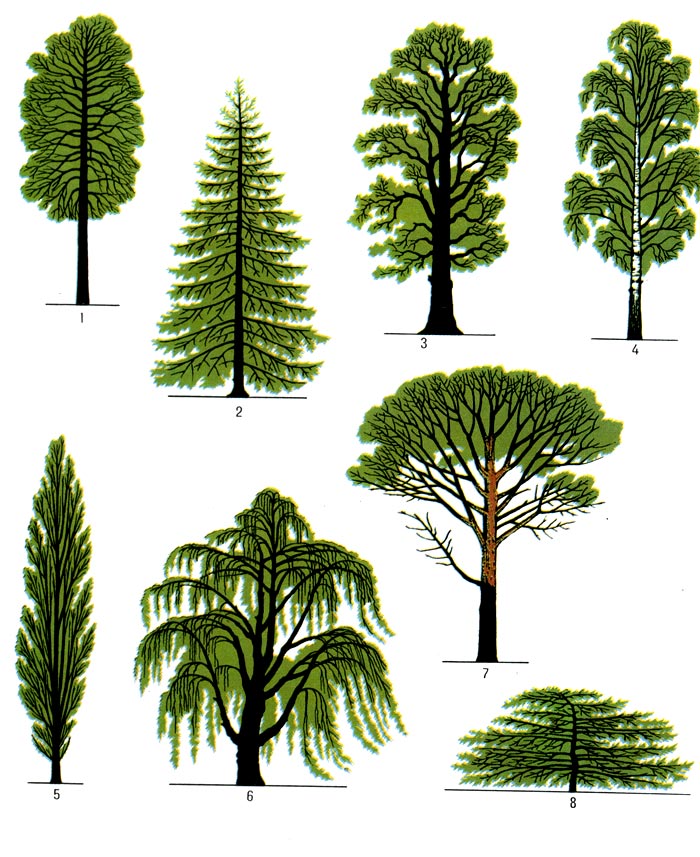 Бывшие ветки, растущие теперь вертикально, превращаются в новые стволы уже со своим собственным густым ветвлением, что происходит за счёт улучшенного питания благодаря новым корням. Эти стволы образуют единую крону.
Бывшие ветки, растущие теперь вертикально, превращаются в новые стволы уже со своим собственным густым ветвлением, что происходит за счёт улучшенного питания благодаря новым корням. Эти стволы образуют единую крону.
Стиль мертвая древесина (сяримики)
По прошествии времени, в результате воздействия суровых погодных условий на стволах некоторых деревьев образуются лысые участки без коры. Обычно они начинаются у основания ствола на уровне грунта, и становятся все более тонкими выше по стволу. Интенсивный солнечный свет отбеливает их, создавая весьма живописные элементы дерева. В бонсай для искусственного создания такого эффекта кора снимается острым ножом, а обнаженная древесина после высыхания отбеливается сернистой известью.
Порядок получения семейного образования
В настоящее время на территории Российской Федерации согласно требованиям действующего законодательства форма получения общего образования и форма обучения по конкретной основной общеобразовательной программе определяются родителями (законными представителями) несовершеннолетнего с учетом его мнения.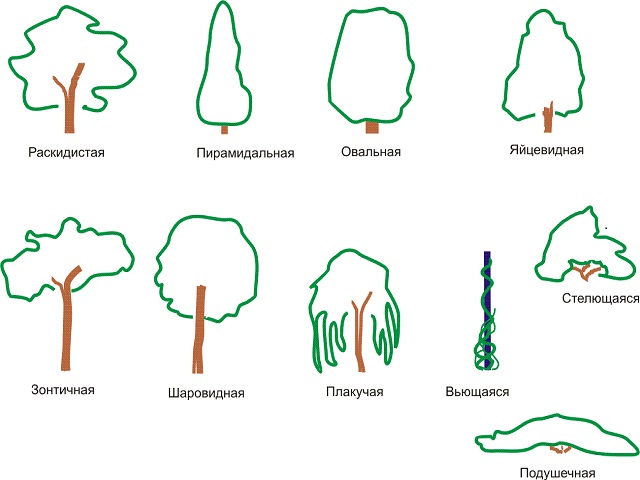
В 2021/2022 учебном году образовательные организации вправе применять новые федеральные государственные образовательные стандарты (ФГОС) начального и основного общего образования для обучения детей, зачисленных до вступления в силу соответствующего приказа, утвердившего ФГОС, с согласия родителей несовершеннолетних и с согласия самих обучающихся.
Семейное обучение — это форма получения образования вне школы. Никаких особых условий для перехода на семейное обучение нет. Переход возможен в любое время.
Основанием (причиной) для перехода на получение семейного образования является возможность при такой форме образования учитывать индивидуальные особенности ребенка, его образовательные потребности, в т. ч. нормировать ежедневные учебные нагрузки, составляя график обучения и режим занятий, выбирать методы подачи материала. Образовательная организация только лишь проверяет знания детей на аттестации.
В случае выбора родителями формы получения общего образования в виде семейного образования они информируют об этом выборе орган местного самоуправления муниципального района или городского округа, на территории которых проживают.
В соответствии с приказом Министерством просвещения Российской Федерации от 22.03.2021 № 115 сроки для подачи уведомления о переходе на семейное обучение: в течение 15 календарных дней с момента утверждения приказа об отчислении обучающегося из образовательной организации в связи с переходом на семейное образование или не менее чем за 15 календарных дней до начала учебного года, в котором планируется переход на семейное образование.
Родители сами выбирают школу для прохождения промежуточной и государственной итоговой аттестации и обращаются в нее с письменным заявлением, в том числе для получения графика прохождения промежуточной аттестации. При этом необязательно выбирать школу по прописке. Порядок проведения аттестации и ее регулярность определяется школой самостоятельно. Учебниками и учебными пособиями обучающиеся обеспечиваются из библиотечного фонда образовательной организации, где будет проходить аттестацию.
При желании на семейном обучении можно участвовать в олимпиадах и конкурсах, а также посещать школьные кружки и секции.
Важно, что при выборе семейного обучения для получения общего образования именно родители обязаны обеспечить целенаправленное овладение обучающимся знаниями, навыками, уметь их применять в повседневной жизни. Кроме того, на родителей возлагается формирование у несовершеннолетнего мотивации получения образования. Образовательная организация в данном случае ответственность не несет.
Также, немаловажно, что обучающийся, получающий образование в форме семейного образования на любом этапе обучения вправе продолжить образование в любой иной форме, предусмотренной законодательством, либо сочетать формы получения образования и обучения.
Яблоня и груша Форма дерева
Несколько веществ стимулируют ветвление у различных сортов яблони и грущи в питомнике [529—536]. Среди них 3-фенил-1,2,4-тиадиазол-5-ил-тиоуксусная кислота, пропил-З-трег-бутилфеиоксиацетат и смесь эфиров жирных кислот. Пропил-З-трег-бутилфенокснацетат хорошо известен по своему сокращенному (кодовому) названию М В 25—105, которое зарегистрировано в качестве фирменного наименования этого вещества [536]. Деревья вишни и сливы также отзывчивы на обработку этим веществом. Другим соединением, которое эффективно стимулирует ветвление у яблони, груши и сливы, является этил-5 (4-хлорфенол) 2Н-тетразол-2-ил-ацетат [537—538]. Обработка этим соединением не только улучшает форму кроны молодых деревьев, но и ускоряет цветение и плодоношение, способствуя при этом увеличению урожая. Результаты, полученные на деревцах черешни, пока не особенно обнадеживают. [c.64]
Деревья вишни и сливы также отзывчивы на обработку этим веществом. Другим соединением, которое эффективно стимулирует ветвление у яблони, груши и сливы, является этил-5 (4-хлорфенол) 2Н-тетразол-2-ил-ацетат [537—538]. Обработка этим соединением не только улучшает форму кроны молодых деревьев, но и ускоряет цветение и плодоношение, способствуя при этом увеличению урожая. Результаты, полученные на деревцах черешни, пока не особенно обнадеживают. [c.64]
Низкоштамбовая форма дерева применяется для выращивания яблони, груши, персика, сливы и вишни. Хотя каждая из этих культур имеет свой вариант низкоштамбовой формы, основные принципы ее обрезки остаются неизменными. Формирование кроны дерева может занять до пяти лет. Цветочные почки, правда, нередко закладываются уже на третий-четвертый год, однако хороший урожай получается обычно еще через год. [c.112]
Карликовая пирамида как одна из форм кроны широко применяется в промышленном садоводстве. Это довольно простой способ интенсивного производства яблок и груш. Низкорослые деревья получают прививкой на карликовые подвои. В частности, груша, выращиваемая на айве, хорошо подходит для этого способа формирования, который в последние годы применяется и для сливы. У яблони и груши формируют дерево с центральным проводником высотой около 2 м и средней длиной веток около 1 м причем крона сужается к верхушке, образуя пирамиду. [c.124]
Это довольно простой способ интенсивного производства яблок и груш. Низкорослые деревья получают прививкой на карликовые подвои. В частности, груша, выращиваемая на айве, хорошо подходит для этого способа формирования, который в последние годы применяется и для сливы. У яблони и груши формируют дерево с центральным проводником высотой около 2 м и средней длиной веток около 1 м причем крона сужается к верхушке, образуя пирамиду. [c.124]
Парша яблони и груши — заболевание, вызываемое грибком, который поражает листья и плоды. Грибок зимует на опавших листьях. Пораженные грибком плоды изменяют свою форму и на них появляются пятна бурого цвета. Зараженные грибком листья опадают. Борьбу с грибком проводят путем опрыскивания деревьев 1%-ным раствором бордосской жидкости и сжигания листьев осенью. [c.96]
Кроме СССР, выпускается в США, ГДР и Японии в форме смачивающегося порошка с содерл анием действующего вещества не менее 60%. У нас выпускается в виде 86% чного смачивающегося порошка, который следует применять в концентрации 0,4— 0,6% по препарату. Против парши яблони и груши первое опрыскивание следует начинать до цветения, второе — в конце цветения, последующие два — через 7—10 дней. В случае распространения парши последующие обработки деревьев надо проводить с интервалами в 10— 14 дней. [c.60]
Против парши яблони и груши первое опрыскивание следует начинать до цветения, второе — в конце цветения, последующие два — через 7—10 дней. В случае распространения парши последующие обработки деревьев надо проводить с интервалами в 10— 14 дней. [c.60]
Яблонная запятовидная щитовка (рис. Ш). Распространена повсеместно. Повреждает яблоню, грушу, сливу, липу, ясень, терн, сирень, боярышник, кизильник и другие породы. Щиток самки вытянутый и довольно часто изогнут наподобие запятой, длиной 2—4 мм, коричневатый, тело грушевидной формы, мо-лочно-белое, длиной до 1,4 мм. Яйца молочно-белые, зимуют под щитками отмерших самок на коре стволов и ветвей. Весной, вскоре после цветения яблони, из яиц отрождаются бледно-желтые личинки, которые присасываются к коре и покрываются щитками. Развитие личинок продолжается 40—50 дней, после чего они превращаются в самок. Последние откладывают по 50— 100 яиц и погибают. Самцы щитовки на плодовых деревьях встречаются редко, этот вид размножается чаще всего без оплодотворения. Развивается в одном поколении. [c.100]
Развивается в одном поколении. [c.100]
Цинковая и марганцевая недостаточность в Новой Зелан- дии установлена для яблони, груши, айвы, вишни и других плодовых культур. Применяется опрыскивание плодовых деревьев растворами сульфата цинка и сульфата марганца с добавлением негашеной извести. Широко используется молибден для улучшения лугов, а также под посевы бобовых трав и овощных культур. Молибден вносят главным образом в виде молибденового суперфосфата, приготавливаемого на заводах путем добавления 1,8 фунта (815 г) молибдата натрия на 1 т суперфосфата. За 1952—1953 гг. в Новой Зеландии было применено 5422 т молибденового суперфосфата, а за первые 4 месяца 1953—1954 гг. применение его увеличилось до 9788 г. Широко распространен недостаток кобальта, наблюдающийся на пемзовых почвах, а также на сильно выщелоченных почвах, образовавшихся на породах вулканического происхождения. Кобальт применяется в форме кобальтового суперфосфата, изготовляемого на заводах путем смешения 3—6 фунтов (1361 — [c. 297]
297]
Калифорнийская щитовка (рис. III). Наиболее опасный вредитель плодовых деревьев. Объект внутреннего карантина растений. Распространена калифорнийская щитовка в Краснодарском и Ставропольском краях, в Волгоградской, Астраханской, Ростовской областях, на Кавказе, Молдавии, Средней Азии, на Дальнем Востоке. Многоядна. Повреждает более 150 видов растений. Щитки самок круглые, диаметром до 2 мм, слегка вы-щ клые, коричневатые, легко отделяются от тела, глаза, ноги и усики отсутствуют. Тело самцов светло-оранжевое, длиной 0,85 мм, с хорошо развитыми усиками, одной парой крьшьев и пурпурно-красными глазами. Личинка первого возраста (бродяжка) продолговато-овальная, уплощенная, от светло-желтой до светло-оранжевой, длиной до 0,25 мм. Личинки второго возраста по форме и окраске тела напоминают взрослую самку, длиной 0,42 мм. Зимуют ушедшие в диапаузу личинки первого возраста и частично взрослые самки. Распространяется калифорнийская щитовка с посадочным и прививочным материалом, плодами, а бродяжки и с помощью ветра.
 На поврежденных побегах и плодах, в местах питания щитовок, образуются красные пятна, листья опадают, плоды, особенно зимних сортов яблони и груши, теряют товарные качества, побеги искривляются, кора растрескивается. При сильном заражении усыхают-отдельные ветви, молодые деревья часто полностью погибают. Калифорнийская щитовка дает несколько поколений в год, каждое из которых развивается около двух месяцев. [c.100]
На поврежденных побегах и плодах, в местах питания щитовок, образуются красные пятна, листья опадают, плоды, особенно зимних сортов яблони и груши, теряют товарные качества, побеги искривляются, кора растрескивается. При сильном заражении усыхают-отдельные ветви, молодые деревья часто полностью погибают. Калифорнийская щитовка дает несколько поколений в год, каждое из которых развивается около двух месяцев. [c.100] Черный рак (рис. VH). Опасное грибное заболевание яблони и в меньшей мере груши. Распространено главным образом в центральных, южных и юго-восточных районах садоводства. Из-за высокой вредоносности черный рак получил название антонов огонь . Часто вызывает гибель деревьев и значительный недобор урожая плодов. Болезнь может поражать кору стволов и ветвей, листья, цветки и плоды. Основная и наиболее опасная форма болезни — поражение коры на штамбах и скелетных ветвей дерева. Первоначально на коре появляются буро-фиолетовые, слегка вдавленные пятна, разрастающиеся концентрическими зоналш. В дальнейшем з частки пораженной коры растрескиваются и чернеют, как бы обугливаются, на них появляется большое количество очень мелких черных бугорков — пикнид со спорами возбудителя заболевания. Кора в местах поражения становится слегка бугристой, напоминая гусиную кожу. Больные участки постепенно расширяются и опоясывают кольцом отмершей ткани ветви и стволы, вызывая неизбежную гибель отдельных ветвей либо всей кроны. Пораженные листья покрываются красновато-коричневыми пятнами, нередко засыхают и опадают. Отмершая ткань на них часто бывает разделена концентрическими кругами. На пораженных плодах появляются бурые вдавленные пятна с чередующимися более светлыми кругами. Полностью побуревшие плоды приобретают темно-бурую или блестяще-черную окраску, сморщиваются, мумифицируются и покрываются многочисленными мелкими бугорками — пикнидами. Возможно также заражение цветков, которые сморщиваются и засыхают. В течение лета болезнь распространяется спорами. Зимуют пикниды со спорами на пораженных участках коры, мумифицированных плодах и опавших листьях.
В дальнейшем з частки пораженной коры растрескиваются и чернеют, как бы обугливаются, на них появляется большое количество очень мелких черных бугорков — пикнид со спорами возбудителя заболевания. Кора в местах поражения становится слегка бугристой, напоминая гусиную кожу. Больные участки постепенно расширяются и опоясывают кольцом отмершей ткани ветви и стволы, вызывая неизбежную гибель отдельных ветвей либо всей кроны. Пораженные листья покрываются красновато-коричневыми пятнами, нередко засыхают и опадают. Отмершая ткань на них часто бывает разделена концентрическими кругами. На пораженных плодах появляются бурые вдавленные пятна с чередующимися более светлыми кругами. Полностью побуревшие плоды приобретают темно-бурую или блестяще-черную окраску, сморщиваются, мумифицируются и покрываются многочисленными мелкими бугорками — пикнидами. Возможно также заражение цветков, которые сморщиваются и засыхают. В течение лета болезнь распространяется спорами. Зимуют пикниды со спорами на пораженных участках коры, мумифицированных плодах и опавших листьях. [c.142]
[c.142]
Плодовые деревья, в том числе и цитрусовые, при недостатке меди заболевают так называемой суховершинностью, или экзантемой. Это заболевание отмечено в ряде стран и проявляется оно обычно на кислых песчаных, а также на некоторых карбонатных почвах. Кроме цитрусовых, этому заболеванию подвержены груша, яблоня, слива, абрикос, персик, маслина и ряд других плодовых культур . Признаки недостатка меди у цитрусовых довольно подробно описаны в работе А. Кемпа, X. Чемпена и Э. Паркера . Авторы указывают, что первым признаком сравнительно слабо выраженной медной недостаточности у цитрусовых служит появление необычно широких темно-зеленых листьев на длинных мягких побегах листья приобретают неправильные очертания с дугообразным выгибо.м средней жилки. Концы побегов прогибаются и частично принимают форму буквы 8. При более сильном голодании листья могут быть очень мелкими и быстро опадают в связи с начит [c.111]
У груши симптомы недостатка цинка появляются в тех же условиях, что и у яблони. Черешня еще более чувствительна к недостатку цинка, чем яблоня, которая в свою очередь более чувствительна по сравнению с другими плодовыми культурами, ореховыми или цитрусовыми. -Признаки цинкового голодания у черешни выражаются в появлении мелких, узких и деформированных хлоротичных листьев. Хлороз вначале появляется на краях листьев и постепенно распространяется к средней жилке листа. При сильном развитии заболевания весь лист постепенно становится желтым или белым . Недостаток цинка у персиковых деревьев характеризуется хлорозной крапчатостью листьев, появляющейся в конце лета. Первыми поражаются нижние листья побегов затем постепенно хлороз распространяется вверх . Признаки цинкового голодания иногда появляются и на однолетних деревьях, но обычно это заболевание отмечается на двухлетних и более старых деревьях. При острой форме цинкового голодания вслед за мелколистностью на следующий год может произойти отмирание ветвей, а в особо тяжелых случаях через 3—4 года может наступить полная гибель дерева.
Черешня еще более чувствительна к недостатку цинка, чем яблоня, которая в свою очередь более чувствительна по сравнению с другими плодовыми культурами, ореховыми или цитрусовыми. -Признаки цинкового голодания у черешни выражаются в появлении мелких, узких и деформированных хлоротичных листьев. Хлороз вначале появляется на краях листьев и постепенно распространяется к средней жилке листа. При сильном развитии заболевания весь лист постепенно становится желтым или белым . Недостаток цинка у персиковых деревьев характеризуется хлорозной крапчатостью листьев, появляющейся в конце лета. Первыми поражаются нижние листья побегов затем постепенно хлороз распространяется вверх . Признаки цинкового голодания иногда появляются и на однолетних деревьях, но обычно это заболевание отмечается на двухлетних и более старых деревьях. При острой форме цинкового голодания вслед за мелколистностью на следующий год может произойти отмирание ветвей, а в особо тяжелых случаях через 3—4 года может наступить полная гибель дерева. [c.237]
[c.237]
При формировании кордона, важно так контролировать рост, чтобы все части дерева находились в пределах вытянутой руки. Поэтому кордон обычно высаживается под углом 45° и имеет один ствол (одноплечий). Но его можно выращивать и вертикально, и в виде многоплечего кордона. Яблоню и грушу, как правило, выращивают в форме наклон- [c.116]
Форма дерева 2.3 APK + Mod (Unlimited money) для Android
Tree Shape — это топиарийная игра. Давайте веселиться в формировании деревьев. Игра Tree Shape — это искусство формирования дерева. Ухаживайте за деревьями и делайте из них разные формы! Вы садовник, и вы должны срезать ветви дерева. У вас есть идеи по дизайну деревьев? Ваша цель — сделать фигуру из большого дерева. Используйте ножницы для обрезки деревьев и нарисуйте дерево красивыми формами.Вы — садовник, лесоруб, портной и парикмахер в мире деревьев. Придумайте идею дизайна дерева и формируйте деревья лучше, чем резчик по дереву. Уход за растениями требует тех же навыков, что и у парикмахера и портного. Играйте в игру Tree Shape и играйте в дерево приключений.Игра топиарий веселее, чем игра в парикмахерскую и портной. Используйте ножницы и покажите свои навыки резки на деревьях. Если вы хорошо подстригаете деревья, то вы играете в Scissorman of Tree Shape.Мы с нетерпением ждем ваших ценных отзывов и предложений.
Уход за растениями требует тех же навыков, что и у парикмахера и портного. Играйте в игру Tree Shape и играйте в дерево приключений.Игра топиарий веселее, чем игра в парикмахерскую и портной. Используйте ножницы и покажите свои навыки резки на деревьях. Если вы хорошо подстригаете деревья, то вы играете в Scissorman of Tree Shape.Мы с нетерпением ждем ваших ценных отзывов и предложений.
Против информации
Свободная покупкаКак установить
Установите шаги:Сначала вы должны удалить оригинальную версию Форма дерева если вы ее установили.
Затем загрузите Форма дерева Мод APK на нашем сайте.
После завершения загрузки вы должны найти файл apk и установить его.
Вы должны включить «Неизвестные источники» для установки приложений за пределами Play Store.
Тогда вы можете открыть и наслаждаться Форма дерева Мод APK
Является Форма дерева Мод Сейф?
Форма дерева Мод на 100% безопасен, потому что приложение было проверено нашей платформой Anti-Malware и вирусы не обнаружены. Антивирусная платформа включает в себя: AOL Active Virus Shield, avast !, AVG, Clam AntiVirus и т. Д. Наш антивирусный движок фильтрует приложения и классифицирует их в соответствии с нашими параметрами. Следовательно, установка на 100% безопасна. Форма дерева Мод APK на нашем сайте.
Антивирусная платформа включает в себя: AOL Active Virus Shield, avast !, AVG, Clam AntiVirus и т. Д. Наш антивирусный движок фильтрует приложения и классифицирует их в соответствии с нашими параметрами. Следовательно, установка на 100% безопасна. Форма дерева Мод APK на нашем сайте.Жизненные формы древесных растений
Жизненные формы древесных растений
Термин «жизненная форма растений» впервые предложил датский ботаник Е. Варминг в 1884 г. Этот термин означает форму, в которой вегетативное тело растения (индивида) находится в гармонии с внешней средой в течение всей его жизни.
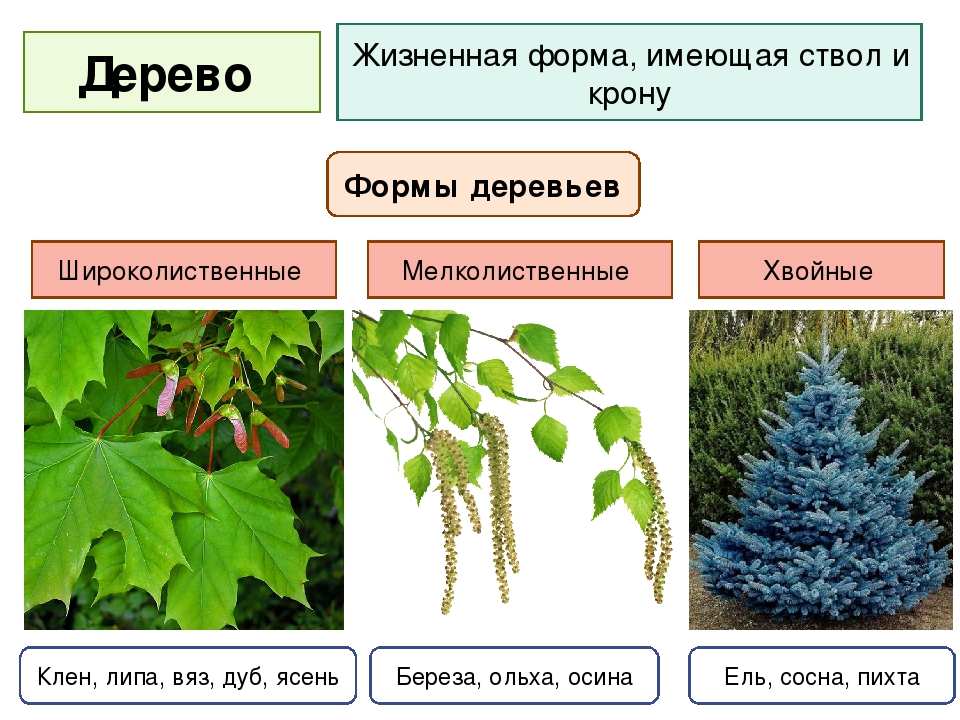
Деревья
Дерево — эволюционно наиболее древний тип жизненной формы семенных растений, возникший около 400 млн. лет назад. Деревья всегда обладают достаточно развитым одревесневшим стволом, разветвленным или неветвящимся, сохраняющимся в течение всей жизни растения — от десятков до тысячи лет.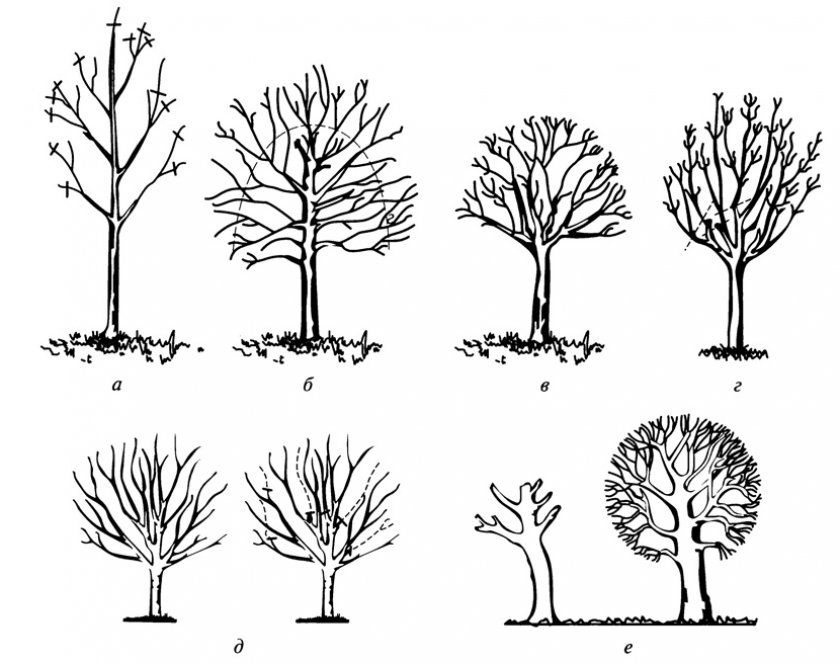 Высота деревьев может составлять от 2-5 до 100 м и более. Деревья включают разные группы жизненных форм. Образователями древесной растительности России являются деревья лесного, кустовидного, лесостепного, сезонно-суккулентного типов и деревья-стланцы.
Высота деревьев может составлять от 2-5 до 100 м и более. Деревья включают разные группы жизненных форм. Образователями древесной растительности России являются деревья лесного, кустовидного, лесостепного, сезонно-суккулентного типов и деревья-стланцы.
Деревья лесного типа
Деревья лесного типа — главные образователи лесов. Их ствол, единственный в течение всего онтогенеза, длительное время сохраняет резкое преобладание по длине и толщине над боковыми ветвями (явление апикального доминирования). Даже в кроне главная ось заметно выделяется по толщине среди боковых ветвей (виды ели, пихты, лиственницы, сосны, дуба, тополя). После рубки или отмирания ствола у многих древесных пород этого типа (секвойя, дуб, бук, вяз, береза) из спящих почек могут вырастать два или несколько вторичных (порослевых) стволов.
Деревья кустовидного типа
Деревья кустовидного типа во взрослом состоянии имеют несколько стволов, развивающихся из спящих (или придаточных) почек у основания материнского ствола. Но в отличие от деревьев лесного типа боковые стволы здесь возникают не в результате удаления материнского ствола, а в связи с его естественным старением. Деревья этого типа (ольха серая, рябина обыкновенная, береза извилистая) представляют собой переходные формы от деревьев к кустарникам.
Но в отличие от деревьев лесного типа боковые стволы здесь возникают не в результате удаления материнского ствола, а в связи с его естественным старением. Деревья этого типа (ольха серая, рябина обыкновенная, береза извилистая) представляют собой переходные формы от деревьев к кустарникам.
Деревья лесостепного, или плодового типа
Деревья лесостепного, или плодового типа характеризуются стволом, рано теряющим преобладание в росте над боковыми ветвями. Поэтому крона начинается близ поверхности почвы, а в самой кроне главная ось не выделяется среди сильных боковых ветвей (виды яблони, абрикоса, сливы, айва обыкновенная, клены татарский и приречный).
Сезонно-суккулентные деревья
Сезонно-суккулентные деревья — обитатели засушливых (аридных) областей, в СНГ — пустынь Средней Азии (саксаул). Из-за сильной редукции листьев практически безлистны (афильные растения). Функции органов ассимиляции у них выполняют зеленые суккулентные однолетние побеги, опадающие в течение жаркого и сухого лета или осени.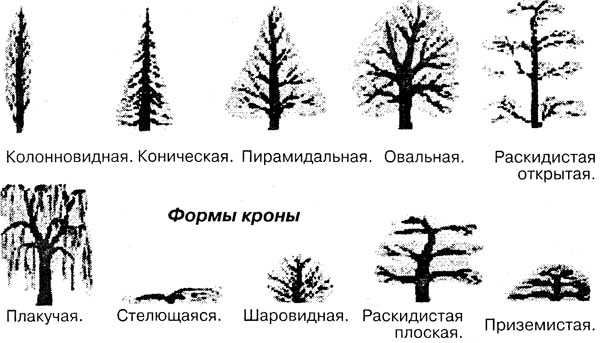 Образователями же кроны являются побеги другого типа: многолетние несуккулентные одревесневающие.
Образователями же кроны являются побеги другого типа: многолетние несуккулентные одревесневающие.
Деревья-стланцы
У деревьев-стланцев главный ствол довольно рано полегает на землю и укореняется. Укореняться способны и скелетные ветви. Деревья этого типа (сосна кедровая стланиковая, можжевельник туркестанский и др.) распространены в субальпийском поясе гор, близ северных границ леса, а иногда на торфяниках и песках в таежной зоне.
Кустарник
У кустарников главный ствол выражен только в первые годы жизни растения. Затем он теряется среди равных ему или даже более мощных надземных стеблей (скелетных осей), последовательно возникающих из спящих почек; позже ствол отмирает. Большинство видов кустарников несет полностью одревесневающие удлиненные побеги. Но есть и суккулентностеблевые (виды кактусов), а также розеточные виды (кустарниковидные пальмы). Среди кустарников с полностью одревесневающими удлиненными побегами различают прямостоячие (виды лещины, барбариса, розы, сирени, жимолости), полупростратные и стелющиеся, у которых главная ось и боковые ветви лежачие, укореняющиеся, но приподнимающиеся у верхушки. Такие кустарники (сосна горная стланиковая, ольховник кустарниковый, горные заросли ивняков и кустарниковых видов можжевельника) широко распространены в субальпийских и субарктических областях, образуя криволесье. В отличие от деревьев длительность жизни надземных скелетных ветвей кустарников в большинстве случаев невелика: 10-20 лет (от 2-3 до 40 лет и более). Высота кустарников от 0,8-1 до 5-6 м, диаметр надземных скелетных осей от 1-2 до 5-8 см.
Такие кустарники (сосна горная стланиковая, ольховник кустарниковый, горные заросли ивняков и кустарниковых видов можжевельника) широко распространены в субальпийских и субарктических областях, образуя криволесье. В отличие от деревьев длительность жизни надземных скелетных ветвей кустарников в большинстве случаев невелика: 10-20 лет (от 2-3 до 40 лет и более). Высота кустарников от 0,8-1 до 5-6 м, диаметр надземных скелетных осей от 1-2 до 5-8 см.
Кустарники широко распространены от экваториальных областей до холодных зон.
Кустарнички
Кустарнички — древесные растения, у которых главная ось имеется лишь в начале онтогенеза. Затем она сменяется боковыми надземными осями, образующимися из спящих почек базальной (нижней, у основания) части материнской оси. Поэтому во взрослом состоянии кустарнички имеют большое число ветвящихся скелетных осей, связанных друг с другом надземно и подземно и последовательно сменяющихся в течение онтогенеза растений. Длительность жизни прямостоячих надземных осей у кустарничков обычно не превышает 5-10 лет, а высота растений может составлять от 5-7 до 50-60 см. Среди кустарничков преобладают вечнозеленые (вереск, брусника, клюква, толокнянка, водяника, линнея), но есть и листопадные (голубика) или такие, как черника: до 10-12 лет она вечнозеленая, а позже становится листопадной. Кустарнички широко распространены в тундре, лесотундре, тайге и высокогорных областях.
Длительность жизни прямостоячих надземных осей у кустарничков обычно не превышает 5-10 лет, а высота растений может составлять от 5-7 до 50-60 см. Среди кустарничков преобладают вечнозеленые (вереск, брусника, клюква, толокнянка, водяника, линнея), но есть и листопадные (голубика) или такие, как черника: до 10-12 лет она вечнозеленая, а позже становится листопадной. Кустарнички широко распространены в тундре, лесотундре, тайге и высокогорных областях.
Полукустарники
Полукустарники — полудревесные растения, у которых удлиненные побеги на значительной части длины ежегодно остаются травянистыми и отмирают. Сохраняются и одревесневают лишь базальные части надземных осей. В отличие от деревьев и кустарников у полукустарников почки возобновления располагаются только близ поверхности почвы. Обитают они преимущественно в засушливых областях (виды полыни, астрагала, тмина, шлемника, дрока). К полукустарникам также принято относить многолетние растения типа малины, ежевики и малиноклена. У них побеги обычно одревесневают полностью, но живут только два года. В первый год побеги несут листья и почки возобновления, во второй — листья, цветки и плоды. После созревания плодов побеги отмирают, а на смену им вырастают новые побеги, опять-таки с двухлетним циклом развития.
У них побеги обычно одревесневают полностью, но живут только два года. В первый год побеги несут листья и почки возобновления, во второй — листья, цветки и плоды. После созревания плодов побеги отмирают, а на смену им вырастают новые побеги, опять-таки с двухлетним циклом развития.
Лианы
Лианы — растения с гибкими неустойчивыми стеблями, которые для своего роста в высоту нуждаются в опоре. Лианы могут быть древовидными (виды гнетума, ротанговых пальм, винограда, актинидии), кустарниковыми со стеблями не толще 10 см (виды древогубца, лимонника, виноградовика), кустарничковыми (плюш, обыкновенный), полукустарниковыми (паслен сладко-горький). Некоторые древовидные лианы являются самыми длинными растениями на Земле; например, отдельные виды ротанговой пальмы способны достигать 300 м в длину.
Древесные растения-подушки
Древесные растения-подушки — жизненная форма, возникающая в крайне жестких условиях существования (пустыня, тундра, высокогорье). Для растений-подушек (виды руты, астрагала, волчеягодника, молочая) характерны ничтожно малый прирост, сильная редукция листьев, выровненная поверхность подушки высотой от 0,1 до 1 м.
Для растений-подушек (виды руты, астрагала, волчеягодника, молочая) характерны ничтожно малый прирост, сильная редукция листьев, выровненная поверхность подушки высотой от 0,1 до 1 м.
Все многообразие жизненных форм растений на Земле отражает как различные уровни приспособленности их к условиям внешней среды, так и разные этапы эволюции.
[Булыгин Н.Е., «Дендрология», 2-е изд., Агропромиздат, 1991.]
Руководство по различным формам деревьев для вашего двора
Выбирая деревья для посадки во дворе, важно выбрать те, форма которых соответствует вашему общему дизайну. Например, если вы хотите что-то высокое и тонкое, вы добьетесь гораздо большего успеха, если выберете дерево, которое естественным образом растет в форме высокой колонны, а не пытается подрезать широкое раскидистое дерево.
Поскольку каждое дерево отличается от других, присвоение названий формам — это нечто меньшее, чем точная наука, но в целом они делятся на категории. Ниже приводится руководство по основным формам обычных ландшафтных деревьев:
Ниже приводится руководство по основным формам обычных ландшафтных деревьев:
Колонновидные деревья высокие и очень тонкие, с вертикальными ветвями. Их вертикальная форма увеличивает высоту, а также обеспечивает отличное экранирование, не занимая много места в ландшафте.
Примеры: Кипарис итальянский, тополь ломбардский, дуб пирамидальный, туя изумрудно-зеленая.
Деревья пирамидальной или конической формы
Мы часто думаем о рождественских елках как о классических пирамидальных деревьях, но эта форма применима и к лиственным деревьям.Пирамидальные деревья шире внизу, с основным центральным стволом и горизонтальными ветвями. Ветви могут начинаться на уровне земли или выше по стволу. Конические деревья похожи, но обычно более тонкие и имеют форму пули. Эти деревья очень эффектны и нуждаются в пространстве, чтобы достичь своей полной ширины.
Примеры: голубая ель, пихта Фрейзера, дуб дуб, западный красный кедр.
Деревья в форме вазы — это именно то, на что они похожи: центральный ствол, который разветвляется в вертикальную дугообразную форму с самой широкой вершиной.Деревья в форме вазы изящны и идеально подходят для облицовки дорожек, потому что они дают тень и пространство над головой.
Примеры: креп мирт, американский вяз, кванзанская вишня.
Деревья круглой или овальной формы
Эти деревья прямостоячие, с сильным центральным стволом, который разветвляется в густую круглую или овальную крону. Они создают сильную тень и могут иметь такую густую листву, что скрывают ветки.
Примеры: клен сахарный, груша Бредфорд, ясень белый, кислица.
Раскидистые или открытые деревья
Эти деревья имеют открытую, неправильную форму, которая может быть в ширину больше, чем в высоту. Некоторые из них представляют собой возвышающиеся над горизонтом деревья, а другие представляют собой небольшие, густые деревья-образцы. Как правило, они создают пятнистую тень и отлично подходят для смягчения зданий.
Как правило, они создают пятнистую тень и отлично подходят для смягчения зданий.
Примеры: вишня, кизил, гинкго, мимоза.
Плакучие деревья
Плакучие деревья имеют гибкие длинные ветви, которые свисают вниз и могут даже касаться земли.Форма этих деревьев часто неправильная и драматичная, что делает их отличным образцом или акцентным деревом. Плакучие деревья не следует сажать возле пешеходных дорожек или улиц, где свисающие ветви будут мешать или их нужно будет обрезать.
Примеры: плакучая ива, плакучая вишня, плакучая шелковица.
Деревья в форме зонтика
Деревья в форме зонтика имеют довольно высокие ветви, которые широко раскинуты, образуя навес зонтика. Если ветви достаточно высоки, эти деревья идеально подходят для затенения зон отдыха или патио, потому что они, естественно, оставляют ясный вид снизу.
Примеры: цветущая вишня, акация, клен японский, пало-верде.
Дополнительная информация
Использование статистики формы филогенетического дерева для определения базовой структуры популяции хозяина | BMC Bioinformatics
Разработка моделей структурированных и неструктурированных популяций хозяев
В этом исследовании мы рассмотрели динамику как структурированных, так и неструктурированных популяций. Структурированная популяция была разбита на две подгруппы, так что люди в этих подгруппах были неразличимы, в то время как между подгруппами существовала разница [34].Выбор двух подпопуляций был поддержан предыдущей работой, в которой две группы использовались для изучения структуры популяции. [3, 17, 35].
Рис. 5Структурированная популяция хозяина с двумя субпопуляциями \(S_1\) и \(S_2\)
Структурированная популяция состояла из субпопуляций \(S_1\) и \(S_2\), как показано на рис. 5. В данной субпопуляции могли произойти только три основных события: рождение, миграция и смерть. Все эти события происходили с разной частотой между субпопуляциями.Скорость, с которой индивидуум в подгруппе \(S_i\) рождал другого в этой подгруппе, был \(\lambda _i\), для \(i \in \{1, 2\}\). Человек в \(S_i\) умер со скоростью \(\mu _i\), для \(i \in \{1, 2\}\). Особь из \(S_i\) мигрировала в \(S_j\) со скоростью миграции \(\alpha _{ij}\), где \(i, j \in \{1, 2\}\).
Все эти события происходили с разной частотой между субпопуляциями.Скорость, с которой индивидуум в подгруппе \(S_i\) рождал другого в этой подгруппе, был \(\lambda _i\), для \(i \in \{1, 2\}\). Человек в \(S_i\) умер со скоростью \(\mu _i\), для \(i \in \{1, 2\}\). Особь из \(S_i\) мигрировала в \(S_j\) со скоростью миграции \(\alpha _{ij}\), где \(i, j \in \{1, 2\}\).
В отличие от структурированной популяции, в неструктурированной популяции все индивидуумы вели себя одинаково. Другими словами, не было субпопуляций, разделяющих неструктурированную популяцию.Моделирование неструктурированной популяции следовало за процессом рождения-смерти без миграционных событий.
Моделирование филогенетических деревьев в структурированных и неструктурированных популяциях
Для моделирования филогенетических деревьев мы учли все возможные события, которые могут произойти во время эволюции популяции. Для структурированной популяции мы позволяем \(N_i(t)\) представлять количество существующих родословных в \(S_i\) в момент времени t для \(i \in \{1, 2\}\).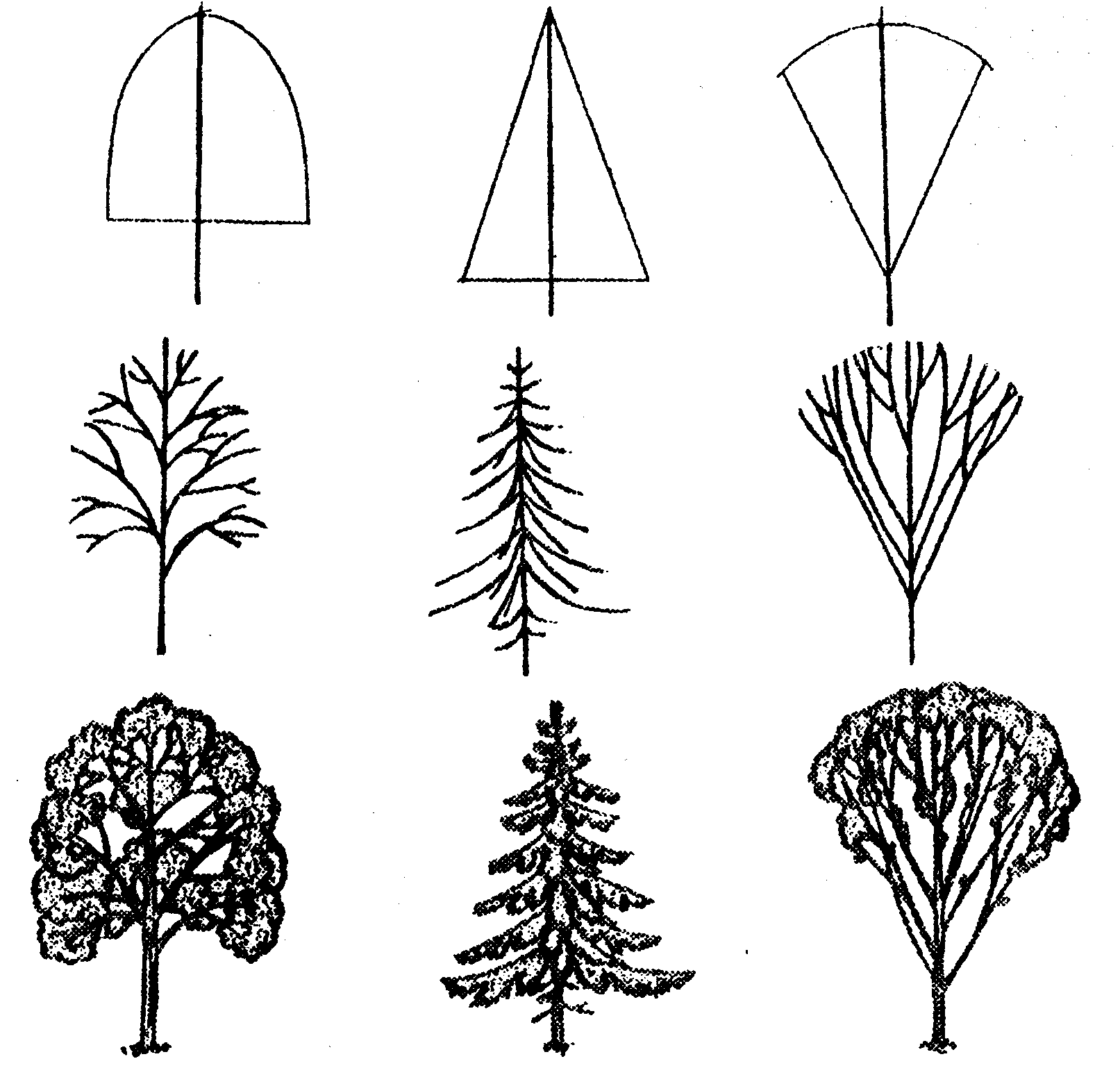 Линия является репрезентативной для одного инфицированного человека.{-N_i(t)(\lambda _i + \mu _i + \alpha _{ij})x} &{} x \ge 0 \\ 0 &{} x < 0. \\ \end{array} \right . \end{aligned}$$
Линия является репрезентативной для одного инфицированного человека.{-N_i(t)(\lambda _i + \mu _i + \alpha _{ij})x} &{} x \ge 0 \\ 0 &{} x < 0. \\ \end{array} \right . \end{aligned}$$
Каждый из наших процессов симуляции дерева начинался с одного человека либо в \(S_1\), либо в \(S_2\). Любое из трех событий (рождение, миграция или смерть) произошло случайным образом.
Чтобы определить подгруппу, в которой произошли продолжающиеся события, мы определили промежуточные показатели в \(S_i\) как \(T_{Si}=\lambda _i + \mu _i + \alpha _{ij}\), для \(i, j \in \{1, 2\}\). Мы случайным образом нарисовали число от 0 до 1, скажем, n .
Если
$$\begin{align} n \le \frac{T_{Si}}{T_{Si} + T_{Sj}}, \end{align}$$
следующее событие произошло в \(S_i\), и если условие не было выполнено, то событие произошло в \(S_j\). Предполагая, что выбранной подгруппой для следующего события является \(S_i\), мы определили следующее событие, вытягивая случайным образом число от 0 до 1, скажем, \(\sigma\).
- (я)
Если
$$\begin{aligned} \sigma \le \frac{\lambda _i}{T_{Si}}, \end{aligned}$$
то событием было рождение.
- (ii)
Если
$$\begin{aligned} \frac{\lambda _i}{T_{Si}} <\sigma \le \frac{\lambda _i + \alpha _{ij}}{T_{Si}}, \end{aligned}$$
, тогда событием была миграция.
- (iii)
В противном случае событие означало смерть.
Моделирование было прекращено после достижения заданного количества существующих родословных. Однако моделирование можно остановить по истечении заданного времени t или возраста дерева. Наша процедура моделирования была реализована в программном обеспечении Python версии 3..jpg) 7.3. Был использован «ETE 3 Toolkit» [36]. Мы смоделировали два набора филогенетических деревьев как для структурированных, так и для неструктурированных популяций.
7.3. Был использован «ETE 3 Toolkit» [36]. Мы смоделировали два набора филогенетических деревьев как для структурированных, так и для неструктурированных популяций.
Включение \(R_0\) в моделирование филогенетического дерева
Базовое репродуктивное число, обозначаемое как \(R_0\), является одним из важных эпидемиологических параметров болезни, который можно оценить непосредственно на основе геномных филогений (Stadler et al.2012) [37]. \(R_0\) измеряет количество вторичных случаев, вызванных одной первичной инфекцией, занесенной во все восприимчивую популяцию [38]. В своей простейшей форме \(R_0\) зависит от частоты контактов, вероятности контакта, вызывающего инфекцию (восприимчивый человек заражается при контакте с инфицированным человеком) и продолжительности инфекционного периода [38]. Для инфекционных заболеваний значение \(R_0>\) 1 связано со вспышкой и персистенцией инфекции [38]. С другой стороны, если \(R_0 \le 1\), то будут реализовываться мелкие вспышки с вероятностью исчезновения единицы [38].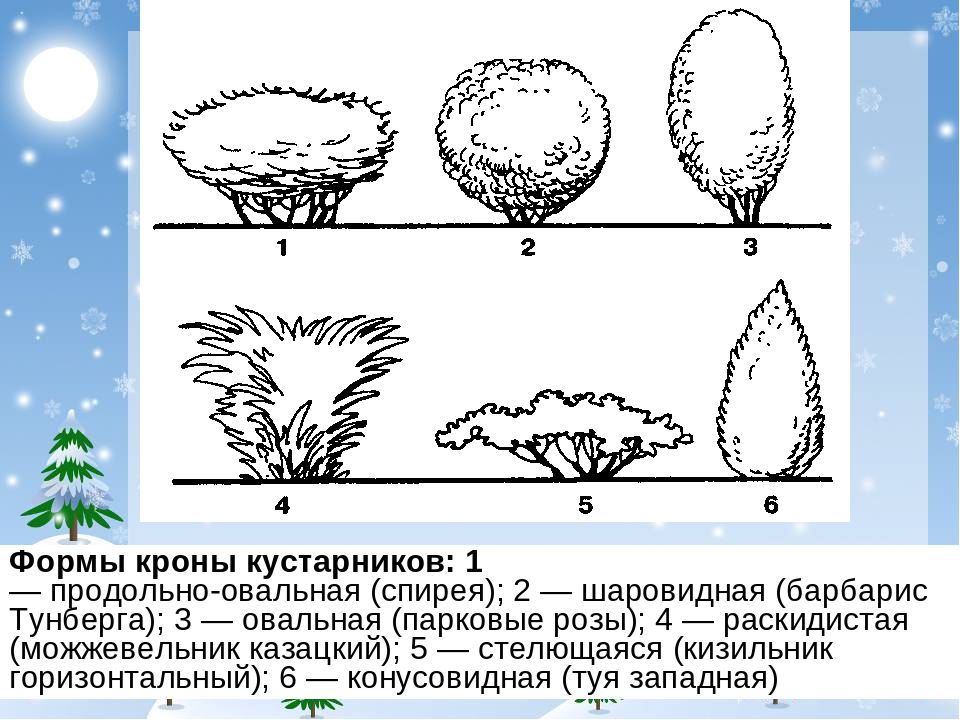
Мы исследовали детерминистский характер модельного процесса рождения-смерти, это было направлено на установление условий, при которых инфекция (процесс) сохраняется. Это осознанный выбор параметров, при которых процесс моделирования был осуществим. Следующая система обыкновенных дифференциальных уравнений описывает динамику инфекции в двух субпопуляциях \(S_1\) и \(S_2\), показанных на рис. 5, с параметрами рождения, смерти и миграции, как описано ранее.
$$\begin{align} \dfrac{dS_1}{dt}&= (\lambda _1S_1 + \alpha _{21}S_2) — (\mu _1S_1 + \alpha _{12}S_1), \end{ выровнено}$$
(1)
$$\begin{aligned} \dfrac{dS_2}{dt}&= (\lambda _2S_2 + \alpha _{12}S_1) — (\mu _2S_2 + \alpha _{21}S_2).*) = (0,0)\) .
Затем мы вычислили \(R_0\) этой системы, используя матричный метод следующего поколения [39, 40]. В отличие от типичных эпидемиологических моделей, оба компартмента репрезентативны для инфекционных заболеваний. Определим X следующим образом:
$$\begin{aligned} X&= \begin{bmatrix} S_1 \\ S_2 \end{bmatrix} \end{aligned}$$
Затем мы построили матрицы \({ \mathcal {F}}\) и \({\mathcal {V}}\) для новых и других терминов заражения в соответствующих разделах:
$$\begin{aligned} {\mathcal {F}}&= \begin{bmatrix} \lambda _1S_1 \\ \lambda _2S_2 \end{bmatrix} = \begin{bmatrix} f_1 \\ f_2 \end{bmatrix} , ~~~~ {\mathcal {V}} = \begin{bmatrix } \mu _1S_1 + \alpha _{12}S_1 — \alpha _{21}S_2 \\ \mu _2S_2 + \alpha _{21}S_2 — \alpha _{12}S_1 \end{bmatrix} = \begin{ bmatrix} v_1 \\ v_2 \end{bmatrix} \end{aligned}$$
Матрицы Якоби \({\mathcal {F}}\) и \({\mathcal {V}}\) в DFE были получено как:
$$\begin{aligned} F&= \begin{bmatrix} \dfrac{\partial f_1}{\partial S_1} &{} \dfrac{\partial f_1}{\partial S_2} \\ \\ \dfrac{\partial f_2}{\partial S_1} &{} \dfrac{\partial f_2}{\partial S_2} \end{bmatrix} _{(0,0)} = \begin{bmatrix} \lambd a _1 &{} 0 \\ 0 &{} \lambda 2 \end{bmatrix}. {-1}\), и получили соответствующие собственные значения.{2} — \alpha _{12} \lambda _{1} \lambda _{2} — \lambda _{1} \lambda _{2} \mu _{1}\right) } \mu _{2 } \end{aligned} \end{aligned}$$
{-1}\), и получили соответствующие собственные значения.{2} — \alpha _{12} \lambda _{1} \lambda _{2} — \lambda _{1} \lambda _{2} \mu _{1}\right) } \mu _{2 } \end{aligned} \end{aligned}$$
Приведенное выше выражение для \(R_0\) применимо к общему случаю структурированной совокупности с двумя подгруппами, \(S_1\) и \(S_2\) . Мы рассмотрели два случая получения \(R_0\), в первом случае мы рассмотрели одну популяцию без субпопуляций. У нас были только рождаемость и смертность без параметров скорости миграции. Таков был сценарий при неструктурированном населении.Во втором случае параметры рождаемости, смертности и миграции в одной субпопуляции умножались на коэффициент k параметров другой субпопуляции. Этот случай представлял собой структурированную популяцию. В обоих случаях мы установили связь между параметрами, для которых \(R_0>1\).
- 1.
\(\lambda _1=\lambda _2=\lambda\), \(\mu _1=\mu _2=\mu\), \(\alpha _{12}=\alpha _{21}=0\) ,
\(R_0 = \dfrac{\lambda }{\mu } ~~~\text {для}~ R_0> 1 \подразумевает \lambda > \mu\).

- 2.
\(\lambda _2=k\lambda _1\), \(\mu _2=k\mu _1\), \(\alpha _{21}=k\alpha _{12}\), где \(k >0\),
\(R_0 = \dfrac{\lambda _1}{2\alpha _{12} + \mu _1}~~\text {for}~~R_0> 1 \подразумевает \lambda _1 > 2\alpha _{12} + \мю _1\).
Выбор параметров, используемых в моделировании филогенетического дерева
Поскольку нас интересовало применение наших методов к реальному набору данных, таких как данные о последовательностях ВИЧ/СПИДа, параметры, которые мы использовали в моделировании филогенетического дерева, были основаны на литературе, связанной с ВИЧ/СПИДом. в Уганде.Что касается уровня смертности, [41] сообщил, что предполагаемая смертность взрослых от ВИЧ/СПИДа составила 21 000 (17 000 29 000) из общего числа 1 500 000 (1 400 000 1 600 000) взрослых, живущих с ВИЧ/СПИДом. Это соответствует коэффициенту смертности 0,014 (0,01 0,02). Поскольку [42] заметил, что распространенность ВИЧ была в три раза выше в сообществах с высоким риском заражения по сравнению с населением в целом, мы умножили на 3, чтобы получить параметр уровня смертности для подгрупп высокого риска в структурированном населении.Для \(R_0\) выбор был основан на работе [43], где мы использовали \(R_0\) 4,99 (0,45, 6,34) для неструктурированного населения и 9,09 (4,18, 36,75) для структурированного населения. Для параметра скорости миграции использовалось значение 0,3 (0,18, 0,44) для подгруппы низкого риска и 0,2 (0,10, 0,33) для подгруппы высокого риска в структурированной популяции. Это было основано на работе [44]. Затем мы вычислили параметр рождаемости на основе формулы для \(R_0\). Значения параметров из литературы, которые имеют отношение к нашему исследованию, показаны в таблице 4.
Это соответствует коэффициенту смертности 0,014 (0,01 0,02). Поскольку [42] заметил, что распространенность ВИЧ была в три раза выше в сообществах с высоким риском заражения по сравнению с населением в целом, мы умножили на 3, чтобы получить параметр уровня смертности для подгрупп высокого риска в структурированном населении.Для \(R_0\) выбор был основан на работе [43], где мы использовали \(R_0\) 4,99 (0,45, 6,34) для неструктурированного населения и 9,09 (4,18, 36,75) для структурированного населения. Для параметра скорости миграции использовалось значение 0,3 (0,18, 0,44) для подгруппы низкого риска и 0,2 (0,10, 0,33) для подгруппы высокого риска в структурированной популяции. Это было основано на работе [44]. Затем мы вычислили параметр рождаемости на основе формулы для \(R_0\). Значения параметров из литературы, которые имеют отношение к нашему исследованию, показаны в таблице 4.
Наборы смоделированных деревьев
Для набора данных 1 (базовый набор данных) : \(\mu =0,014, R_0=4,99, \lambda =\mu (R_0)\), количество вершин было 350, а количество деревьев — 500. Для структурированной совокупности в подгруппе 1 (низкий риск) использовались следующие параметры: \(\mu _1=0.014,~\alpha _{12}=0,3,~R_{01}=4,99,~\lambda _1=R_{01}(2\alpha _{12}+\mu _1)\), число наконечников составило 350, а количество деревьев 250. Для подгруппы 2 (высокий риск) использовались следующие параметры: \(\mu _2=3(0,014),~\alpha _{21}=0,2,~R_{02} =9,09,~\lambda _2=R_{02}(2\alpha _{21}+\mu _2)\), количество подсказок 200 и количество деревьев 250.
Для структурированной совокупности в подгруппе 1 (низкий риск) использовались следующие параметры: \(\mu _1=0.014,~\alpha _{12}=0,3,~R_{01}=4,99,~\lambda _1=R_{01}(2\alpha _{12}+\mu _1)\), число наконечников составило 350, а количество деревьев 250. Для подгруппы 2 (высокий риск) использовались следующие параметры: \(\mu _2=3(0,014),~\alpha _{21}=0,2,~R_{02} =9,09,~\lambda _2=R_{02}(2\alpha _{21}+\mu _2)\), количество подсказок 200 и количество деревьев 250.
Для набора данных 2 количество наконечников варьировали, сохраняя при этом другие значения параметров для набора данных 1 постоянными. Для структурированной популяции количество подсказок варьировалось в интервале (300, 400) для субпопуляции 1 и (150, 250) для субпопуляции 2.Для неструктурированной популяции советы варьировались в интервале (300, 400). Сводка значений параметров, использованных для набора данных 1, показана в таблице 4.
Оценка статистики деревьев по смоделированным деревьям
Формы деревьев, смоделированных как по структурированным, так и по неструктурированным популяциям, были исследованы с использованием оценок статистики деревьев. Эти статистические данные о деревьях включали: количество вишен; индексы Сакина, Коллесса и суммарные кофенетические индексы; длина лестницы; максимальная ширина; индекс отношения максимальной глубины и ширины к глубине.Подробное описание количества вишен было сделано [18, 29, 32]. Для индекса Сакина подробности можно найти в работах [18, 24]. Индекс Коллесса был определен и описан в [24, 45]. Суммарный кофенетический индекс — один из новых индексов для филогенетических деревьев, определенный в работе [33]. Определения и описания для длины лестницы, максимальной глубины дерева, максимальной ширины дерева и индекса отношения ширины к глубине были сделаны [25]. Сводка определений статистики восьми деревьев показана в таблице 5.На рис. 1 показано смоделированное дерево с 5 подсказками, а в таблице 6 показано, как вычислялась статистика соответствующего дерева. Все вычисления для восьми статистических данных были реализованы в программном обеспечении Python версии 3.7.3.
Эти статистические данные о деревьях включали: количество вишен; индексы Сакина, Коллесса и суммарные кофенетические индексы; длина лестницы; максимальная ширина; индекс отношения максимальной глубины и ширины к глубине.Подробное описание количества вишен было сделано [18, 29, 32]. Для индекса Сакина подробности можно найти в работах [18, 24]. Индекс Коллесса был определен и описан в [24, 45]. Суммарный кофенетический индекс — один из новых индексов для филогенетических деревьев, определенный в работе [33]. Определения и описания для длины лестницы, максимальной глубины дерева, максимальной ширины дерева и индекса отношения ширины к глубине были сделаны [25]. Сводка определений статистики восьми деревьев показана в таблице 5.На рис. 1 показано смоделированное дерево с 5 подсказками, а в таблице 6 показано, как вычислялась статистика соответствующего дерева. Все вычисления для восьми статистических данных были реализованы в программном обеспечении Python версии 3.7.3.
 Определения статистики дерева Таблица 6. Вычисленная статистика дерева из дерева, показанного на рис. 1
Определения статистики дерева Таблица 6. Вычисленная статистика дерева из дерева, показанного на рис. 1 Сравнение статистики дерева, рассчитанной для структурированных и неструктурированных совокупностей Различают структурированные и неструктурированные популяции хозяев.Статистика дерева визуализировалась с использованием блочных диаграмм для обеих популяций. Они были полезны для обобщения местоположения и разброса статистики деревьев.
Поскольку ящичковые диаграммы не могли дать четкого различия в статистике дерева между популяциями, мы сначала сравнили распределения статистики дерева для обеих популяций, используя двухвыборочный критерий Колмогорова-Смирнова [47]. Для данной статистики дерева мы исследовали, различает ли этот тест соответствующие распределения в обеих популяциях.Из вычисленных значений p критерий Колмогорова-Смирнова различал деревья как из структурированной, так и из неструктурированной популяции на уровне 0,05. Для этого теста параметр D представляет собой статистику Колмогорова-Смирнова, которая измеряет расстояние между двумя сравниваемыми распределениями. Чем больше параметр D , тем меньше значение p и тем более отдалены (различны) два сравниваемых распределения. Кроме того, мы использовали два непараметрических теста, в том числе тесты Куккони и Подгора-Гаствирта.Это также тесты с двумя выборками, которые определяют, различаются ли две базовые выборки, используя параметры местоположения и масштаба. Эти два и некоторые другие описаны в [48]. Коробчатые диаграммы, двухвыборочные тесты Колмогорова-Смирнова, Куккони и Подгора-Гаствирта были реализованы в R версии 4.0.2. Пакет R «ggplot2» [49] использовался для рисования коробчатых диаграмм.
Для этого теста параметр D представляет собой статистику Колмогорова-Смирнова, которая измеряет расстояние между двумя сравниваемыми распределениями. Чем больше параметр D , тем меньше значение p и тем более отдалены (различны) два сравниваемых распределения. Кроме того, мы использовали два непараметрических теста, в том числе тесты Куккони и Подгора-Гаствирта.Это также тесты с двумя выборками, которые определяют, различаются ли две базовые выборки, используя параметры местоположения и масштаба. Эти два и некоторые другие описаны в [48]. Коробчатые диаграммы, двухвыборочные тесты Колмогорова-Смирнова, Куккони и Подгора-Гаствирта были реализованы в R версии 4.0.2. Пакет R «ggplot2» [49] использовался для рисования коробчатых диаграмм.
Классификация смоделированных деревьев из структурированной или неструктурированной популяции на основе их статистики деревьев
Мы использовали различные алгоритмы классификации для определения типа популяции (структурированной или неструктурированной), из которой был получен данный набор деревьев смоделированные на основе их оценочной статистики деревьев. Алгоритмы классификации, которые мы использовали, были; K-ближайший сосед (KNN) [50], метод опорных векторов (SVM) [51, 52] и деревья решений (DT) [53]. В таблице 7 приведены описания этих классификаторов.
Алгоритмы классификации, которые мы использовали, были; K-ближайший сосед (KNN) [50], метод опорных векторов (SVM) [51, 52] и деревья решений (DT) [53]. В таблице 7 приведены описания этих классификаторов.
Здесь мы стремились разделить смоделированные деревья на два основных класса; , то есть , структурированное и неструктурированное население. Чтобы установить пропорцию, которая будет использоваться для обучения, мы опробовали разные пропорции данных; то есть , \(30\%\), \(50\%\) и \(70\%\) для обучения и с соответствующими оставшимися пропорциями для тестирования классификаторов.Затем мы выполнили поиск по сетке параметров, чтобы получить оптимальные модели, которые будут использоваться для классификации. Процедура классификации оптимальных моделей для KNN, SVM и DT реализована в программе R версии 4.0.2. В классификации использовался R-пакет «e107» [54].
Оценка и перекрестная проверка древовидных классификаторов
Мы сравнили эффективность всех трех методов классификации, используя такие показатели, как чувствительность, специфичность и уровень точности [55].![]() Мы также оценили производительность классификаторов, используя кривые рабочих характеристик приемника (ROC).Для кривой ROC истинные положительные результаты (чувствительность) нанесены на график против ложных положительных результатов (1-специфичность). Вычисляли площадь под кривой (AUC) для ROC-кривых. Некоторый анализ и сравнение ROC-кривых дан в [56]. В нашем исследовании AUC количественно определяет общую способность классификатора различать деревья, смоделированные из структурированных и неструктурированных популяций. Как сообщает [57], AUC колеблется от 0,5 до 1 для реалистичного классификатора. Совершенный классификатор (тот, который имеет ноль ложных срабатываний и ноль ложноотрицательных результатов) имеет площадь 1.Чем ближе значение AUC к 1, тем выше производительность классификатора.
Мы также оценили производительность классификаторов, используя кривые рабочих характеристик приемника (ROC).Для кривой ROC истинные положительные результаты (чувствительность) нанесены на график против ложных положительных результатов (1-специфичность). Вычисляли площадь под кривой (AUC) для ROC-кривых. Некоторый анализ и сравнение ROC-кривых дан в [56]. В нашем исследовании AUC количественно определяет общую способность классификатора различать деревья, смоделированные из структурированных и неструктурированных популяций. Как сообщает [57], AUC колеблется от 0,5 до 1 для реалистичного классификатора. Совершенный классификатор (тот, который имеет ноль ложных срабатываний и ноль ложноотрицательных результатов) имеет площадь 1.Чем ближе значение AUC к 1, тем выше производительность классификатора.
Мы провели 10-кратную перекрестную проверку. При этом данный набор данных делится на 10 равных частей, каждая из которых используется в качестве набора для тестирования, а остальные 9 используются для обучения модели. Затем мы получили средние значения показателей, которые мы использовали для оценки классификаторов. Перекрестная проверка дает реалистичную оценку производительности модели, как это делается в некоторых исследованиях [58, 59]. Настройка параметров модели и перекрестная проверка были выполнены с помощью R-пакета «caret» [60].Оценка и перекрестная проверка древовидных классификаторов были реализованы в программном обеспечении R версии 4.0.2.
Затем мы получили средние значения показателей, которые мы использовали для оценки классификаторов. Перекрестная проверка дает реалистичную оценку производительности модели, как это делается в некоторых исследованиях [58, 59]. Настройка параметров модели и перекрестная проверка были выполнены с помощью R-пакета «caret» [60].Оценка и перекрестная проверка древовидных классификаторов были реализованы в программном обеспечении R версии 4.0.2.
Анализ чувствительности
Чтобы определить, устойчивы ли классификаторы к различному выбору параметров модели и размеру деревьев, мы смоделировали три набора из 250 структурированных и 250 неструктурированных деревьев (всего 500) с (i) случайно выбранными параметры, (ii) случайный размер дерева и (iii) случайные параметры и случайный размер дерева. Параметры и размеры деревьев были получены с использованием выборки латинского гиперкуба, реализованной в «инструментарии» Python «SMT».Параметры были выбраны таким образом, чтобы \(R_0\) находилось в интервалах (0,45, 6,34) и (4,18, 36,75) для субпопуляции 1 и 2 соответственно. Точно так же размер дерева варьировался, чтобы лежать в интервалах (300, 400) и (250, 300) для подпопуляции 1 и 2 соответственно. Выбранные параметры и размеры деревьев были аналогичны реальным популяциям, которые позже были исследованы. Затем смоделированные деревья были классифицированы в соответствии с 10-кратной перекрестной проверкой.
Точно так же размер дерева варьировался, чтобы лежать в интервалах (300, 400) и (250, 300) для подпопуляции 1 и 2 соответственно. Выбранные параметры и размеры деревьев были аналогичны реальным популяциям, которые позже были исследованы. Затем смоделированные деревья были классифицированы в соответствии с 10-кратной перекрестной проверкой.
Применение процедуры классификации к реальным данным
Чтобы оценить эффективность классификаторов на реальных данных в рамках известной эпидемиологии, мы использовали классификаторы на данных о последовательности двух ключевых групп населения в Уганде, чья основная динамика передачи ВИЧ-1 ранее был описан в [61].Мы применили наши классификаторы к филогенетическим деревьям из ранее опубликованных данных о последовательностях ВИЧ \ (-1 \) от населения в целом (GP) и рыбацких сообществ (FC) Уганды, Bbosa et al. [61]. Последовательности были извлечены из базы данных нуклеотидов NCBI, регистрационные номера от MG434786 до MG435347. Данные состояли из 357 последовательностей из GP и 221 последовательности из FC. Были сгенерированы два набора деревьев; (i) только GP (ii) только FC и (iii) комбинация GP и FC. Последовательности были выровнены с использованием «clustalw» [62] перед созданием 1000 бутстрепов деревьев максимального правдоподобия с использованием IQ-TREE Nguyen et al.[63] с UFBoot2 Hoang et al. [64]. Классификаторы SVM, DT и KNN, обученные 10-кратной перекрестной проверке на исходном уровне, использовались для прогнозирования структуры популяции бутстрапов. Мы классифицировали 1000 бутстрапов деревьев с максимальной вероятностью, сгенерированных из трех наборов данных, включая; последовательности из GP, FC и как GP, так и FC.
Были сгенерированы два набора деревьев; (i) только GP (ii) только FC и (iii) комбинация GP и FC. Последовательности были выровнены с использованием «clustalw» [62] перед созданием 1000 бутстрепов деревьев максимального правдоподобия с использованием IQ-TREE Nguyen et al.[63] с UFBoot2 Hoang et al. [64]. Классификаторы SVM, DT и KNN, обученные 10-кратной перекрестной проверке на исходном уровне, использовались для прогнозирования структуры популяции бутстрапов. Мы классифицировали 1000 бутстрапов деревьев с максимальной вероятностью, сгенерированных из трех наборов данных, включая; последовательности из GP, FC и как GP, так и FC.
Форма дерева | Забытые Королевства Вики
Форма дерева Форма дерева , также называемая просто дерево , было заклинанием трансмутации или изменения, которое позволяло заклинателю принимать форму большого дерева или куста на несколько часов. [1] [7] [8] [9]
[1] [7] [8] [9]
Эффекты[]
При применении заклинатель и вся одежда и снаряжение заклинателя превращаются в живое дерево или куст, или мертвый ствол с несколькими мертвыми конечностями. Находясь в форме дерева, заклинатель не мог двигаться, но мог видеть и слышать все, что происходило в непосредственной близости, как если бы заклинатель стоял там обычно. Трансформация прошла все мирские тесты и самую тщательную проверку, но обнаружение магии зарегистрировала слабую магию трансмутации. [1] [7] [8] [9]
Древовидная форма может быть мгновенно убрана заклинателем до истечения срока действия заклинания, без времени восстановления или длительных вредных эффектов. [1] [7] [8] [9]
Трансмутационная версия этого заклинания давала заклинателю дополнительную естественную броню и делала невозможным нанесение одного удара.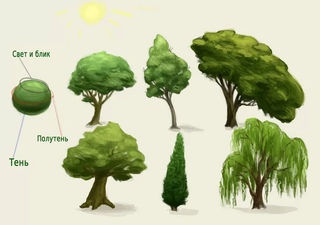 критическое повреждение. [1]
критическое повреждение. [1]
Компоненты[]
Более новая версия древовидной формы требовала использования заклинателем священного символа или божественного фокуса, а также обычных вербальных и соматических компонентов. [1] Для более старой версии модификации также требовалась ветка дерева. [7] [8] [9]
Ссылки[]
- ↑ 1.0 1.1 1.2 1.3 1.4 1.5 Джонатан Твит, Монте Кук, Скип Уильямс). Справочник игрока v.3.5 . (Волшебники побережья), с. 296. ISBN 0-7869-2886-7.
- ↑ Дэвид «Зеб» Кук (август 1989 г.). Справочник игрока (2-е издание) .(TSR, Inc.), стр. 29, 215. ISBN 0-88038-716-5.
- ↑ Дэвид «Зеб» Кук (апрель 1995 г.). Справочник игрока 2-е издание (исправленное) . (TSR, Inc.), стр. 41, 273. ISBN 0-7869-0329-5.
- ↑ Кук, Финдли, Херринг, Кубасик, Сарджент, Лебедь (1991). Фолиант магии 2-е издание . (TSR, Inc), с. 153. ISBN 1-56076-107-5.
- ↑ Ричард Бейкер (1996). Выбор игрока: заклинания и магия . (TSR, Inc), с. 187. ISBN 0-7869-0394-5.
- ↑ Гэри Гайгакс (1978). Справочник игрока 1-е издание . (TSR, Inc.), стр. 25, 58. ISBN 0-9356-9601-6.
- ↑ 7,0 7,1 7,2 7,3 Дэвид «Зеб» Кук (август 1989 г.). Справочник игрока (2-е издание) . (TSR, Inc.), с. 215. ISBN 0-88038-716-5.
- ↑ 8,0 8,1 8,2 8,3 Дэвид «Зеб» Кук (апрель 1995 г.). Справочник игрока 2-е издание (исправленное) . (TSR, Inc.), с. 273. ISBN 0-7869-0329-5.
- ↑ 9,0 9,1 9,2 9,3 Гэри Гайгакс (1978). Справочник игрока 1-е издание . (TSR, Inc.), с. 58. ISBN 0-9356-9601-6.
 Фолиант магии 2-е издание . (TSR, Inc), с. 153. ISBN 1-56076-107-5.
Фолиант магии 2-е издание . (TSR, Inc), с. 153. ISBN 1-56076-107-5.Лесов | Бесплатный полнотекстовый | Изменчивость формы деревьев в смешанном дубовом лесу с использованием наземной лазерной технологии: последствия для анализа системы спаривания
1.
 Введение Изменчивость формы деревьев, особенно форма кроны, может варьироваться между видами деревьев и внутри них в зависимости от многих факторов, на которые влияет место условия и конкуренция деревьев за свет [1].Он также определяется расширением боковых и терминальных ветвей, при этом боковые ветви, отходящие от главного ствола (ветви первого порядка), в той или иной степени контролируются концевыми ветвями. Ветви второго порядка, которые вырастают из первого порядка, контролируются родительской ветвью и так далее. Обычно бывает не более пяти порядков ветвей, так как ветви более высоких порядков отмирают из-за недостатка света внутри кроны [2,3,4]. С точки зрения экологии, расположение ветвей деревьев представляет собой оптимизированную адаптацию к местным условиям [5].Биофизические процессы, связанные с хранением углерода и воды — фотосинтез и эвапотранспирация — также находятся под влиянием архитектуры дерева [6,7]. Архитектура дерева также имеет решающее значение для опыления и спаривания видов, опыляемых ветром.
Введение Изменчивость формы деревьев, особенно форма кроны, может варьироваться между видами деревьев и внутри них в зависимости от многих факторов, на которые влияет место условия и конкуренция деревьев за свет [1].Он также определяется расширением боковых и терминальных ветвей, при этом боковые ветви, отходящие от главного ствола (ветви первого порядка), в той или иной степени контролируются концевыми ветвями. Ветви второго порядка, которые вырастают из первого порядка, контролируются родительской ветвью и так далее. Обычно бывает не более пяти порядков ветвей, так как ветви более высоких порядков отмирают из-за недостатка света внутри кроны [2,3,4]. С точки зрения экологии, расположение ветвей деревьев представляет собой оптимизированную адаптацию к местным условиям [5].Биофизические процессы, связанные с хранением углерода и воды — фотосинтез и эвапотранспирация — также находятся под влиянием архитектуры дерева [6,7]. Архитектура дерева также имеет решающее значение для опыления и спаривания видов, опыляемых ветром. С точки зрения архитектуры дерева важны точность и уровень характеристики, поскольку чем глубже характеристика, тем сложнее и больше времени требуется для извлечения точных результатов. Наземное сканирование LiDAR (TLS) оказалось подходящим методом для оценки дерева. морфологии объективно [8] и недеструктивно [9].TLS является эффективным инструментом, используемым для установления древесной структуры дерева [5,10,11]. Совсем недавно было показано, что TLS может идентифицировать высококачественные характеристики деревьев, которые не поддаются непосредственному измерению в традиционных инвентаризациях леса, такие как объем ствола и компоненты биомассы (общий ствол и ветви). Этот тип входных данных составляет основу точного моделирования, такого как национальные аллометрические модели. Благодаря этому последнему технологическому прогрессу TLS продемонстрировала возможность улучшения качества и количества справочных данных, собранных при инвентаризации леса [12].Следует отметить, что TLS является популярным инструментом в экологии леса для измерения индекса площади листа и кривой ствола.
С точки зрения архитектуры дерева важны точность и уровень характеристики, поскольку чем глубже характеристика, тем сложнее и больше времени требуется для извлечения точных результатов. Наземное сканирование LiDAR (TLS) оказалось подходящим методом для оценки дерева. морфологии объективно [8] и недеструктивно [9].TLS является эффективным инструментом, используемым для установления древесной структуры дерева [5,10,11]. Совсем недавно было показано, что TLS может идентифицировать высококачественные характеристики деревьев, которые не поддаются непосредственному измерению в традиционных инвентаризациях леса, такие как объем ствола и компоненты биомассы (общий ствол и ветви). Этот тип входных данных составляет основу точного моделирования, такого как национальные аллометрические модели. Благодаря этому последнему технологическому прогрессу TLS продемонстрировала возможность улучшения качества и количества справочных данных, собранных при инвентаризации леса [12].Следует отметить, что TLS является популярным инструментом в экологии леса для измерения индекса площади листа и кривой ствола. На внешний вид дерева (то есть на фенотип) влияют как факторы окружающей среды, так и генетическая структура [19]. Гетерозиготность — это мера генетического разнообразия, которая пропорциональна количеству генетической изменчивости в конкретной области генома. Гетерозиготность на индивидуальном уровне использовалась в нескольких случаях для проверки связи между генетическим составом, оцениваемым различными типами генетических маркеров, и характеристиками дерева (например, устойчивостью к загрязнению воздуха [20], характеристиками ствола и кроны [21]. ).Углы ветвей северного красного дуба, по-видимому, подвержены генетическому влиянию [21]. Характеристики ствола и кроны могут объяснить случаи спаривания в лесных насаждениях, выявленные с помощью анализа отцовства и происхождения на основе высокополиморфных ДНК-маркеров. До сих пор только несколько характеристик, описывающих фенотип дерева (например, диаметр на высоте груди, диаметр кроны и объем), были оценены и учтены в исследованиях системы спаривания у лесных древесных пород [22,23].
На внешний вид дерева (то есть на фенотип) влияют как факторы окружающей среды, так и генетическая структура [19]. Гетерозиготность — это мера генетического разнообразия, которая пропорциональна количеству генетической изменчивости в конкретной области генома. Гетерозиготность на индивидуальном уровне использовалась в нескольких случаях для проверки связи между генетическим составом, оцениваемым различными типами генетических маркеров, и характеристиками дерева (например, устойчивостью к загрязнению воздуха [20], характеристиками ствола и кроны [21]. ).Углы ветвей северного красного дуба, по-видимому, подвержены генетическому влиянию [21]. Характеристики ствола и кроны могут объяснить случаи спаривания в лесных насаждениях, выявленные с помощью анализа отцовства и происхождения на основе высокополиморфных ДНК-маркеров. До сих пор только несколько характеристик, описывающих фенотип дерева (например, диаметр на высоте груди, диаметр кроны и объем), были оценены и учтены в исследованиях системы спаривания у лесных древесных пород [22,23]. Система спаривания оказывает важное влияние на количество и распределение генетической изменчивости в популяциях и между ними [24].Помимо размера и плотности популяции [25], на систему спаривания могут влиять несколько факторов, таких как способ опыления и доступность переносчиков [26], синхронизация цветения [27] и степень генетической структуризации популяции [28]. ]. Дубы (Quercus spp.) распространены в северном полушарии и являются важными элементами многих лесных экосистем. Дубы встречаются в самых разных средах, от влажного до более сухого климата. Дубы однодомны, опыляются ветром и имеют тяжелые семена, которые разносятся под действием силы тяжести или птиц (например, птиц).г., сойка) [29,30,31]. В Европе дубы являются одним из доминирующих широколиственных видов умеренного пояса, имеющим важное экономическое, экологическое и социальное значение. Леса, в которых встречается больше видов дуба в симпатрии, предлагают очень хорошие возможности для сравнительного анализа различных аспектов их биологии, включая форму дерева и другие морфологические характеристики.
Система спаривания оказывает важное влияние на количество и распределение генетической изменчивости в популяциях и между ними [24].Помимо размера и плотности популяции [25], на систему спаривания могут влиять несколько факторов, таких как способ опыления и доступность переносчиков [26], синхронизация цветения [27] и степень генетической структуризации популяции [28]. ]. Дубы (Quercus spp.) распространены в северном полушарии и являются важными элементами многих лесных экосистем. Дубы встречаются в самых разных средах, от влажного до более сухого климата. Дубы однодомны, опыляются ветром и имеют тяжелые семена, которые разносятся под действием силы тяжести или птиц (например, птиц).г., сойка) [29,30,31]. В Европе дубы являются одним из доминирующих широколиственных видов умеренного пояса, имеющим важное экономическое, экологическое и социальное значение. Леса, в которых встречается больше видов дуба в симпатрии, предлагают очень хорошие возможности для сравнительного анализа различных аспектов их биологии, включая форму дерева и другие морфологические характеристики. Основная цель настоящего исследования состояла в том, чтобы проанализировать изменчивость формы деревьев различных пород европейского дуба белого, произрастающих в сходных условиях участка.Мы также проверили связь между формой дерева и генетическим составом (то есть индивидуальной гетерозиготностью), а также проверили, имеют ли дубы, идентифицированные как доноры пыльцы в предыдущем исследовании [32], больший размер с точки зрения характеристик кроны и ствола, чем недоноры.
Основная цель настоящего исследования состояла в том, чтобы проанализировать изменчивость формы деревьев различных пород европейского дуба белого, произрастающих в сходных условиях участка.Мы также проверили связь между формой дерева и генетическим составом (то есть индивидуальной гетерозиготностью), а также проверили, имеют ли дубы, идентифицированные как доноры пыльцы в предыдущем исследовании [32], больший размер с точки зрения характеристик кроны и ствола, чем недоноры.4. Обсуждение
Архитектура, развившаяся у отдельного дерева в течение жизненного цикла, является естественной реакцией его реакции на специфические условия местности, особенно с точки зрения конкуренции, доступа к источникам света и контроля положения в условиях гравитации.Понимание трехмерной формы древесной биомассы деревьев является решающим фактором при оценке среды лесопосадки и оценке взаимозависимости между деревьями с точки зрения конкуренции и спаривания; однако при использовании обычных методов получение этих данных занимает много времени и регулярно наносит ущерб (например, полевые оценки с использованием деструктивных методов).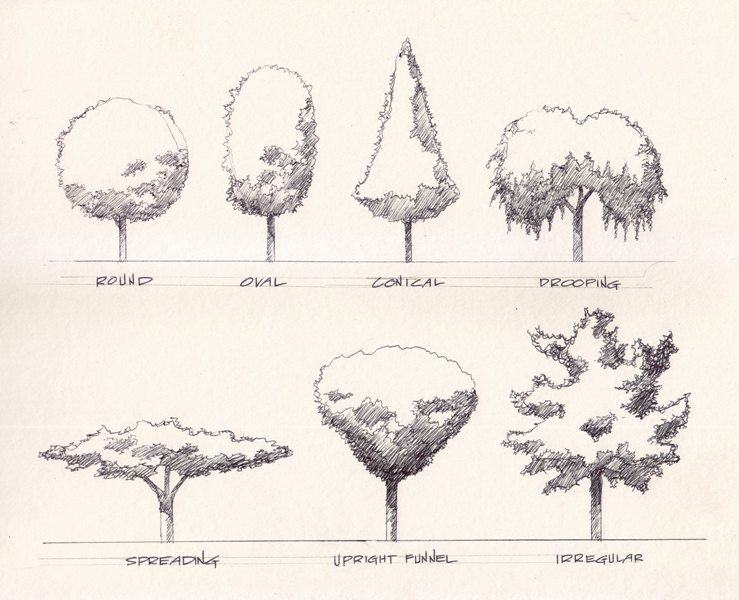 Измерения, которые могут быть достигнуты, особенно с помощью наземного сканера LiDAR (TLS), являются быстрыми, практичными, полными и точными при оцифровке отдельных деревьев или целых участков.TLS также усложнила оценку архитектуры стоящих деревьев в лесных экосистемах с точными оценками объема, биомассы или распределения размеров ветвей [15,16,36]. TLS кажется многообещающей стратегией в лесных средах, особенно при реконструкции архитектуры отдельных деревьев, которые требуют постоянного уровня данных о структуре сети ветвей дерева [5,37].
Измерения, которые могут быть достигнуты, особенно с помощью наземного сканера LiDAR (TLS), являются быстрыми, практичными, полными и точными при оцифровке отдельных деревьев или целых участков.TLS также усложнила оценку архитектуры стоящих деревьев в лесных экосистемах с точными оценками объема, биомассы или распределения размеров ветвей [15,16,36]. TLS кажется многообещающей стратегией в лесных средах, особенно при реконструкции архитектуры отдельных деревьев, которые требуют постоянного уровня данных о структуре сети ветвей дерева [5,37]. Мы получили высокоточные плотные облака точек, которые позволили нам реконструировать ответвления до уровня 7, используя инновационный подход, сочетающий наземные измерения (например,(например, соответствующее расстояние и планируемое расположение станций сканирования), оптимальные погодные условия (например, отсутствие ветра, постоянное освещение) и соответствующее состояние лесной среды (например, отсутствие листвы). Облака точек, полученные с помощью TLS, предоставили дополнительную информацию для различения пород дуба на основе характеристик ствола и кроны. Мы обнаружили значительные различия между различными характеристиками деревьев у всех четырех видов дуба либо на стволе, высоте, либо на уровне ветвей.
Мы обнаружили значительные различия между различными характеристиками деревьев у всех четырех видов дуба либо на стволе, высоте, либо на уровне ветвей.
 , в некодирующей части генов в случае геномных SSR). У сосны обыкновенной не обнаружено связи между гетерозиготностью и характеристиками приспособленности [39]. Однако степень гетерозиготности значительно различалась между клонами ели европейской, отобранными из семенного сада, и случайно выбранными деревьями в лесонасаждениях [40]. Более крупные деревья могут иметь больше мужских сережек и более эффективно рассеивать пыльцу. Мы обнаружили различия в характеристиках кроны и ствола между донорами и недонорами пыльцы, идентифицированными в Бежане путем исключения отцовства с использованием высокополиморфных маркеров ДНК [32].Доноры пыльцы имели больший объем кроны и ветвей, большее количество ветвей и более длинные ветви. Более того, не было существенной разницы в высоте между отобранными дубами, что позволяет предположить, что все отобранные деревья в Бежане имеют одинаковую доступность света. Один показатель размера дерева, DBH, сравнивался с мужской плодовитостью у нескольких видов деревьев [22,23].
, в некодирующей части генов в случае геномных SSR). У сосны обыкновенной не обнаружено связи между гетерозиготностью и характеристиками приспособленности [39]. Однако степень гетерозиготности значительно различалась между клонами ели европейской, отобранными из семенного сада, и случайно выбранными деревьями в лесонасаждениях [40]. Более крупные деревья могут иметь больше мужских сережек и более эффективно рассеивать пыльцу. Мы обнаружили различия в характеристиках кроны и ствола между донорами и недонорами пыльцы, идентифицированными в Бежане путем исключения отцовства с использованием высокополиморфных маркеров ДНК [32].Доноры пыльцы имели больший объем кроны и ветвей, большее количество ветвей и более длинные ветви. Более того, не было существенной разницы в высоте между отобранными дубами, что позволяет предположить, что все отобранные деревья в Бежане имеют одинаковую доступность света. Один показатель размера дерева, DBH, сравнивался с мужской плодовитостью у нескольких видов деревьев [22,23]. . Было обнаружено, что DBH не является важным фактором, влияющим на успех спаривания самцов, что согласуется с нашими выводами. Однако положительная корреляция между DBH и долей оплодотворений наблюдалась в одной популяции Fagus crenata [41].Тем не менее, характеристики кроны, такие как объем и длина ветвей, могут быть более важными для производства пыльцы, чем DBH, которую легко измерить в полевых условиях. Действительно, взаимосвязь между объемом кроны и эффективной мужской плодовитостью была исследована в древостое североамериканского дуба [22]. Слабая, но значимая положительная корреляция была обнаружена между объемом кроны, оцененным по диаметру и высоте кроны, и успешностью мужского оплодотворения дубов. Слабая, но недостоверная положительная связь между числом опылений и объемом ветвей наблюдалась у Q.petraea в лесу Бежан. Отсутствие значимости может быть объяснено низким числом эффективных опылений, обнаруженных в смешанном дубовом лесу методом исключения отцовства [32]. Желуди были собраны в безмачтовый год, поскольку у местных видов дуба интервал между последовательными мачтовыми годами может составлять 10 и более лет [42].
. Было обнаружено, что DBH не является важным фактором, влияющим на успех спаривания самцов, что согласуется с нашими выводами. Однако положительная корреляция между DBH и долей оплодотворений наблюдалась в одной популяции Fagus crenata [41].Тем не менее, характеристики кроны, такие как объем и длина ветвей, могут быть более важными для производства пыльцы, чем DBH, которую легко измерить в полевых условиях. Действительно, взаимосвязь между объемом кроны и эффективной мужской плодовитостью была исследована в древостое североамериканского дуба [22]. Слабая, но значимая положительная корреляция была обнаружена между объемом кроны, оцененным по диаметру и высоте кроны, и успешностью мужского оплодотворения дубов. Слабая, но недостоверная положительная связь между числом опылений и объемом ветвей наблюдалась у Q.petraea в лесу Бежан. Отсутствие значимости может быть объяснено низким числом эффективных опылений, обнаруженных в смешанном дубовом лесу методом исключения отцовства [32]. Желуди были собраны в безмачтовый год, поскольку у местных видов дуба интервал между последовательными мачтовыми годами может составлять 10 и более лет [42].
Диаметр ствола дерева обычно уменьшается или сужается от основания к кончику.То, как происходит это уменьшение, определяет форму штамба. Это сужение может происходить с разной скоростью и разными способами или формами. Понимание формы штамба может позволить:
Существует сложное взаимодействие между формой штамба и кроной дерева. | |
| Геометрические фигуры | Конус геометрической формы правильного очертания можно описать скоростью (k) и формой (b) в общем уравнении: где y обозначает диаметр или радиус, а x обозначает расстояние от вершины формы. Конкретные значения b соответствуют обычным формам:
|
Части ствола дерева имеют тенденцию приближаться к усеченным частям этих общих форм.Основание дерева имеет тенденцию быть неиловидным, а верхушка — конусовидной. Основная часть ствола имеет параболоидную форму. Однако точки перегиба между этими формами не являются постоянными. Формы параболоидов можно дополнительно разбить на квадратичные и кубические параболоиды. Квадратичный параболоид (b = 1) будет генерировать прямую линию, если высота будет построена по отношению к квадрату радиуса, в то время как кубический параболоид (b = 0,66) будет генерировать прямую линию, если высота будет построена по отношению к радиусу в кубе.Мецгер предположил, что ствол дерева должен быть подобен кубическому параболоиду. Он утверждал, что стержень представляет собой балку с равномерным сопротивлением изгибу, закрепленную у основания и функционирующую как плечо рычага. Горизонтальная сила, действующая на такую балку, будет оказывать натяжение, увеличивающееся по направлению к точке крепления, и наиболее экономичной формой для этой балки будет равномерная конусность, подобная усеченному кубическому параболоиду. Грей (1956) (цит. по Carron 1968) утверждал, что ствол дерева не прочно прикреплен к земле.Квадратичная форма параболоида больше соответствовала бы механическим требованиям, налагаемым этим предположением. Приведенные выше механические теории формы ствола являются одним из подходов к объяснению формы дерева. Две другие теории связывают форму ствола дерева с потребностью дерева в транспортировке воды или питательных веществ внутри дерева ( теория проводящей воды и теория питания формы ствола соответственно). Эти теории основаны на идеях, касающихся движения жидкостей по трубам. Гормональная теория стеблевой формы предполагает, что ростовые вещества, образующиеся в кроне, распределяются вокруг и вниз по штамбу, чтобы контролировать активность камбия. Эти вещества будут уменьшать или усиливать радиальный рост в определенных местах штамба и, таким образом, влиять на форму штамба. | |
| Фактор формы | Форм-фактор — это сводная информация об общей форме штока.Объем стебля сравнивается с объемом стандартного геометрического тела того же диаметра в основании и полной высоты. Наиболее распространенным форм-фактором является форм-фактор по высоте груди . Наиболее распространенным форм-фактором является форм-фактор по высоте груди .Стандартная геометрическая форма для форм-фактора высоты груди представляет собой цилиндр той же высоты, что и ножка, и с площадью сечения, равной площади сечения ножки на высоте груди (т. е. базальной площади). Фактор формы – это отношение объема штока к объему цилиндра. Конкретные форм-факторы высоты груди указывают на общие формы ствола:
Если подходящий форм-фактор высоты груди для дерева данного возраста, вида и места можно легко определить, то объем ствола легко вычислить, умножив форм-фактор на высоту дерева и площадь основания. |
| Частное формы | Коэффициент формы представляет собой отношение диаметра в некоторой точке выше уровня груди к диаметру на уровне груди. Опять же, это краткое изложение общей формы ствола. Опять же, это краткое изложение общей формы ствола.Абсолютный коэффициент формы обычно использовался для группировки деревьев в классы форм. Этот коэффициент рассчитывается путем измерения диаметра на высоте, находящейся на полпути между высотой груди и общей высотой дерева.Затем этот диаметр делится на диаметр на уровне груди и выражается в виде десятичной дроби. Коэффициенты абсолютной формы также указывают на общие формы стебля:
Льюис и др. (1976) используют разницу в диаметре между 4,5 и 7 м.5 м и 1,5 м для оценки общей формы Pinus radiata на плантациях Южной Австралии. Это значение используется для улучшения оценки объема ствола. |
| Уравнение конусности Коническая модель | Уравнения конусности разработаны для прогнозирования диаметра в любой точке ствола дерева, т. е. для моделирования профиля ствола. В идеале эти прогнозы непредвзяты и основаны на простых входных переменных, таких как диаметр на высоте груди и общая высота дерева. е. для моделирования профиля ствола. В идеале эти прогнозы непредвзяты и основаны на простых входных переменных, таких как диаметр на высоте груди и общая высота дерева.Построение этих уравнений продолжалось много лет и использовалось множество подходов. Ранние исследователи (например, Fries and Matern, 1965) использовали новые возможности цифровых компьютеров для составления сложных полиномиальных уравнений. Часто ствол дерева делился на две или три части и для каждой части подгонялись отдельные уравнения (например, Max and Burkhart, 1976). Другие исследователи (например, Goulding and Murray, 1975) подчеркивали необходимость использования уравнений конусности для получения оценок, совместимых с предсказаниями объема ствола.Тем не менее, другие подходы (например, Kozak 1988 и Bi 2000) пытаются разработать модель, описывающую непрерывное изменение формы стебля от основания к кончику. Различные подходы к уравнению конусности пытаются преодолеть некоторые общие недостатки:
Например, кубическое полиномиальное уравнение, подобранное для подмножества данных Pinus radiata высота: диаметр из Crook (1998), объяснило 98% вариаций.Общая ошибка в предсказании диаметра была близка к нулю, но были локальные ошибки в культе (в среднем на 1 см занижение диаметра) и на 10-40% ниже высоты штамба (в среднем на 1 см завышено). Эти ошибки были уравновешены соответствующими ошибками выше по стволу. Однако большая часть объема ствола и ценных изделий из древесины, как правило, приходится на нижнюю треть ствола, поэтому эти локальные отклонения значительны. Grosenbaugh (1966) писал: Многие специалисты по измерениям пытались найти единственную простую функцию с двумя переменными, включающую всего несколько параметров, которую можно было бы использовать для описания всего профиля дерева.К сожалению, деревья, по-видимому, способны принимать бесконечное разнообразие форм, и полиномы (или частные полиномы) со степенью по крайней мере на два больше, чем наблюдаемое число перегибов, необходимы для определения различных форм перегибов.Риверс (1995) обнаружил, что локальные погрешности в прогнозировании диаметра могут вызывать значительные погрешности, когда прогнозируемый профиль ствола используется для оценки размера или ценности продуктов на дереве.  В эксперименте с использованием более 100 деревьев он обнаружил, что использование уравнения конусности привело к завышению оценки объема продукта на 10% и завышению стоимости продукта на 14% — уравнение конусности было несмещенным для общего диаметра. Небольшое смещение в оценке диаметра может привести к большой ошибке в значении, если это влияет на предсказание ограничения продукта (например, малого диаметра конца бревна). В эксперименте с использованием более 100 деревьев он обнаружил, что использование уравнения конусности привело к завышению оценки объема продукта на 10% и завышению стоимости продукта на 14% — уравнение конусности было несмещенным для общего диаметра. Небольшое смещение в оценке диаметра может привести к большой ошибке в значении, если это влияет на предсказание ограничения продукта (например, малого диаметра конца бревна). Bi (2000) опубликовал коэффициенты тригонометрического уравнения ленты переменной формы для 26 распространенных видов эвкалипта.Он провел обширные тесты локализованных смещений этой модели. |
[shape.htm] Редакция: 6/1999 |
 Таким образом, любой фактор, влияющий на крону, может влиять и на форму штамба. Различные части штамба растут с разной скоростью, поскольку окружающая среда и другие факторы влияют на крону и способ распределения фотосинтатов.
Таким образом, любой фактор, влияющий на крону, может влиять и на форму штамба. Различные части штамба растут с разной скоростью, поскольку окружающая среда и другие факторы влияют на крону и способ распределения фотосинтатов.

Как сформировать грушу | Главная Руководства
Владелец грушевого дерева должен решать две очень разные проблемы, когда дело доходит до обрезки этого дерева. Эстетическая сторона вопроса крутится вокруг того, как должно выглядеть дерево как во время цветения, так и после, когда оно усыпано зелеными листьями. Другая сторона чисто практическая и сосредоточена на максимизации производства фруктов и хранении всех этих груш в пределах досягаемости.
Другая сторона чисто практическая и сосредоточена на максимизации производства фруктов и хранении всех этих груш в пределах досягаемости.
Отмечайте ветки деревьев для удаления на зиму, проверяя их по определенным критериям, начиная с любой ветки ниже 20 дюймов от земли. Они слишком низкие и должны быть удалены.
Отметьте поврежденные ветки деревьев для удаления, а также любые ветки, не растущие под углом 60-75 градусов. Последние структурно слабы и должны быть удалены, прежде чем они станут проблемой.
Также запланируйте удаление любых перекрывающихся ветвей деревьев. Если смотреть сверху, ветки на груше должны располагаться, как спицы в колесе или часы в часах. Их вертикальное положение (выше или ниже на стволе) не так важно, как их положение на 360-градусной дуге вокруг дерева. Если две конечности уложены одна над другой или почти так, пометьте одну из них ручкой или привяжите к ней ленту.
Отпилите все помеченные ветки от дерева, спилив в точке чуть выше ствола. Не пилите заподлицо с поверхностью ствола дерева.
Не пилите заподлицо с поверхностью ствола дерева.
Проредить внутреннюю часть дерева путем обрезки или отпиливания боковых ветвей от основных ветвей дерева. Оставьте от пяти до восьми веток на ветку.
Отойдите назад и осмотрите дерево целиком. Грушевые деревья обычно имеют яйцевидную или коническую форму, в зависимости от конкретного типа, но должны быть примерно симметричными. Обрежьте любую ветку дерева, которая заметно длиннее аналогичных веток примерно на том же вертикальном уровне, с помощью пилы или ножниц, чтобы восстановить симметрию.
Обрежьте ветки, тянущиеся вверх, если вы хотите, чтобы все дерево оставалось в пределах досягаемости. Если вы используете стремянку для сбора фруктов, обрежьте ветки до удобной высоты стремянки.
Определение эволюционного процесса по форме филогенетического дерева в JSTOR
Абстрактный Выводы о макроэволюционных процессах традиционно зависели исключительно от летописи окаменелостей, но такие выводы можно подкрепить, если также рассмотреть формы филогенетических деревьев, связывающих существующие таксоны. Осознание того, что филогении отражают макроэволюционные процессы, привело к росту литературы, посвященной теоретическим и сравнительным исследованиям формы деревьев. Два аспекта формы дерева особенно важны: баланс дерева и распределение длины ветвей. Мы изучаем и оцениваем последние разработки и связи между этими двумя аспектами и предлагаем направления для будущих исследований. Изучение формы деревьев обещает полезные и мощные проверки макроэволюционных гипотез. При соответствующих дальнейших исследованиях форма дерева может помочь нам обнаруживать массовые вымирания и адаптивную радиацию, измерять непрерывные изменения в темпах видообразования и вымирания и связывать изменения в этих темпах с экологическими или биогеографическими причинами.Полезность формы дерева выходит далеко за рамки изучения макроэволюции. Мы обсуждаем приложения к другим областям биологии, включая коэволюцию, филогенетический вывод, популяционную биологию и биологию развития.
Осознание того, что филогении отражают макроэволюционные процессы, привело к росту литературы, посвященной теоретическим и сравнительным исследованиям формы деревьев. Два аспекта формы дерева особенно важны: баланс дерева и распределение длины ветвей. Мы изучаем и оцениваем последние разработки и связи между этими двумя аспектами и предлагаем направления для будущих исследований. Изучение формы деревьев обещает полезные и мощные проверки макроэволюционных гипотез. При соответствующих дальнейших исследованиях форма дерева может помочь нам обнаруживать массовые вымирания и адаптивную радиацию, измерять непрерывные изменения в темпах видообразования и вымирания и связывать изменения в этих темпах с экологическими или биогеографическими причинами.Полезность формы дерева выходит далеко за рамки изучения макроэволюции. Мы обсуждаем приложения к другим областям биологии, включая коэволюцию, филогенетический вывод, популяционную биологию и биологию развития.
Текущие выпуски теперь доступны на веб-сайте Chicago Journals. Прочитайте последний выпуск. Ежеквартальный обзор биологии (QRB) представляет проницательные исторические, философские и технические трактовки важных биологических тем с 1926 года.Являясь ведущим обзорным журналом по биологии, QRB публикует выдающиеся обзорные статьи большого объема, основанные на обширном, всеобъемлющем и часто гуманистическом понимании биологии. Помимо основных биологических наук, QRB также является важным обзорным журналом для ученых в смежных областях, включая политические исследования, историю и философию науки. Обширный раздел обзоров новых книг по биологии предоставляет преподавателям и исследователям информацию о последних публикациях в жизни
Прочитайте последний выпуск. Ежеквартальный обзор биологии (QRB) представляет проницательные исторические, философские и технические трактовки важных биологических тем с 1926 года.Являясь ведущим обзорным журналом по биологии, QRB публикует выдающиеся обзорные статьи большого объема, основанные на обширном, всеобъемлющем и часто гуманистическом понимании биологии. Помимо основных биологических наук, QRB также является важным обзорным журналом для ученых в смежных областях, включая политические исследования, историю и философию науки. Обширный раздел обзоров новых книг по биологии предоставляет преподавателям и исследователям информацию о последних публикациях в жизни
С момента своего основания в 1890 году в качестве одного из трех основных подразделений Чикагского университета издательство University of Chicago Press взяло на себя обязательство распространять научные знания самого высокого уровня и публиковать серьезные работы, которые способствуют образованию, способствуют общественному пониманию.