
Почему змеи такие разноцветные? Похоже, все дело в печени
Автор фото, Getty Images
Подпись к фото,Змеи дадут людям сто очков вперед в том, что касается оттенков кожи
Ученые из Университета Женевы приблизились к пониманию механизма, который обеспечивает огромное разнообразие расцветки рептилий. Оказалось, что на окрас кожи змей и других пресмыкающихся влияет мутация гена, связанного с очистительной функцией печени.
Чтобы понять природу открытия швейцарских ученых, опубликовавших об этом статью в научном журнале PNAS (официальном издании Национальной академии наук США), нужно понять, как именно змеи — и люди — формируют свою окраску.
Буйство красок
Змеи, рыбы и рептилии выгодно отличаются от остальных позвоночных разнообразием цвета кожи. Если исключить бегающую и летающую позвоночную живность, покрытую шерстью или перьями, то с голой кожей остаются только люди, которых природа обилием красок не побаловала.
Причина яркой и разнообразной палитры змей, равно как и довольно скучной цветовой гаммы Homo sapiens, заключается в хроматофорах, клетках, содержащих пигменты и светоотражающие кристаллы. У рептилий и земноводных их довольно много, причем каждый тип отвечает за свою цветовую гамму.
Автор фото, Ignacio Palacios
Подпись к фото,Хвост хамелеона. Они могут даже перегонять свои хроматофоры из одной части тела в другую, в зависимости от обстановки
Например, меланофоры отвечают за черный и коричневый цвета, ксантофоры — за желтый, эритрофоры — за красный, а отражающие свет иридиофоры придают коже ярко-зеленый или ярко-синий цвета.
В отличие от холоднокровных, у теплокровных птиц, зверей и людей есть только один тип окрашивающих клеток: меланоциты, которые являются до некоторой степени аналогом меланофоров.
Но почему в пределах одного вида пресмыкающихся появляются особи разного цвета? Ответ именно на этот вопрос позволил ученым узнать больше о самом механизме формирования ярких цветов рептилий.
Оказалось, что все дело в мутации гена, связанного с функцией печени.
Эволюционный генетик из Женевского университета Афанасия Цика и ее докторант Асье Юлат-Аготе провели исследование на примере маисовых полозов.
Автор фото, Mike Wesemann
Подпись к фото,Яркий маисовый полоз…
Эта небольшая неядовитая змея, обитающая на юге Северной Америки, известна большим разнообразием окраса. Ее еще называют красной крысиной змеей, или гутатой.
Есть как минимум семь самых распространенных вариантов окраса (их называют морфами) маисовых полозов — от ярко-оранжевого с черной окантовкой до бледно-розового и нежно фиолетового. Последний морф считается для биологов самым интересным из-за почти полного отсутствия меланофоров. Таких пастельно окрашенных змей называют «лавандовыми полозами».
Автор фото, LA Dawson
Подпись к фото,… и его бледный родственник
При чем тут печень?
Согласно выводам швейцарских ученых, рождение нежно-розовых полозов вызвано мутацией одного-единственного гена (LYST, или CHS1), отвечающего за выработку специфического белка, который тоже называется LYST.
Автор фото, MesserWoland and Szczepan1990
Подпись к фото,Структура клетки. Лизосома — под номером 12
Эта аббревиатура расшифровывается как «Регулятор лизосомального транспорта». Этот белок отвечает за перенос различных молекул в лизосомы, компоненты клеток-гепатоцитов, работающих как микрозаводы по переработке мусора.
Но какое отношение имеют лизосомы к окраске маисовых полозов?
Ученые из Женевского университета проанализировали гепатоциты, основные клетки печени всех позвоночных. У них множество разнообразных функций, в том числе они отвечают за вывод из организма вредных и опасных веществ.
Автор фото, Alexquaas
Подпись к фото,Клетки печени под электронным микроскопом. Гепатоциты составляют от 60 до 80 процентов ее веса
У розовых полозов лизосомы гепатоцитов оказались гораздо большими — и они более активно и кардинально меняли структуру хроматофоров, отвечающих за яркие цвета.
Как отметила Афанасия Цика, исследование было долгим и кропотливым, ведь змеи откладывают яйца всего раз в год, и чтобы выявить нужный ген, ученым пришлось изучить весь геном маисового полоза.
Однако изучение всех причин разнообразия в окрасе рептилий и рыб еще продолжается.
К чему приводит мутация гена LYST у людей
Автор фото, Brocken Inaglory
Подпись к фото,Альбинизмом страдают не только люди
Ген LYST имеется и в человеческом организме. У людей его мутация вызывает так называемый Синдром Чедиака-Хигаси, который приводит к альбинизму (отсутствию пигмента меланина) и опасному ослаблению иммунной системы.
На настоящий момент это заболевание неизлечимо, хотя в некоторых случаях помогает пересадка костного мозга.
что такое в Ударении и правописании
Смотреть что такое КУКУРУЗНЫЙ в других словарях:
КУКУРУЗНЫЙ
кукурузный прил. 1) Соотносящийся по знач. с сущ.: кукуруза (1), связанный с ним. 2) Свойственный кукурузе (1), характерный для нее. 3) Предназначенный для уборки, обработки кукурузы (1). 4) Сделанный, изготовленный из кукурузы (2).<br><br><br>… смотреть
с сущ.: кукуруза (1), связанный с ним. 2) Свойственный кукурузе (1), характерный для нее. 3) Предназначенный для уборки, обработки кукурузы (1). 4) Сделанный, изготовленный из кукурузы (2).<br><br><br>… смотреть
КУКУРУЗНЫЙ
кукурузный прил. к кукурузакукурузная мука — corn meal
КУКУРУЗНЫЙ
кукурузный маисовый Словарь русских синонимов. кукурузный прил., кол-во синонимов: 6 • вердепешевый (4) • детской неожиданности (3) • кукурузовый (1) • маисовый (1) • пшеничковый (1) • цвет (266) Словарь синонимов ASIS.В.Н. Тришин.2013. . Синонимы: вердепешевый, детской неожиданности, кукурузовый, маисовый, цвет… смотреть
КУКУРУЗНЫЙ
1) Орфографическая запись слова: кукурузный2) Ударение в слове: кукур`узный3) Деление слова на слоги (перенос слова): кукурузный4) Фонетическая транскр… смотреть
КУКУРУЗНЫЙ
кукуру́зный,
кукуру́зная,
кукуру́зное,
кукуру́зные,
кукуру́зного,
кукуру́зной,
кукуру́зного,
кукуру́зных,
кукуру́зному,
кукуру́зной,
кукуру́зному,
кукуру́зным,
кукуру́зный,
кукуру́зную,
кукуру́зное,
кукуру́зные,
кукуру́зного,
кукуру́зную,
кукуру́зное,
кукуру́зных,
кукуру́зным,
кукуру́зной,
кукуру́зною,
кукуру́зным,
кукуру́зными,
кукуру́зном,
кукуру́зной,
кукуру́зном,
кукуру́зных,
кукуру́зен,
кукуру́зна,
кукуру́зно,
кукуру́зны,
кукуру́знее,
покукуру́знее,
кукуру́зней,
покукуру́зней
(Источник: «Полная акцентуированная парадигма по А. А. Зализняку»)
.
Синонимы:
вердепешевый, детской неожиданности, кукурузовый, маисовый, цвет… смотреть
А. Зализняку»)
.
Синонимы:
вердепешевый, детской неожиданности, кукурузовый, маисовый, цвет… смотреть
КУКУРУЗНЫЙ
кукуру’зный, кукуру’зная, кукуру’зное, кукуру’зные, кукуру’зного, кукуру’зной, кукуру’зного, кукуру’зных, кукуру’зному, кукуру’зной, кукуру’зному, кукуру’зным, кукуру’зный, кукуру’зную, кукуру’зное, кукуру’зные, кукуру’зного, кукуру’зную, кукуру’зное, кукуру’зных, кукуру’зным, кукуру’зной, кукуру’зною, кукуру’зным, кукуру’зными, кукуру’зном, кукуру’зной, кукуру’зном, кукуру’зных, кукуру’зен, кукуру’зна, кукуру’зно, кукуру’зны, кукуру’знее, покукуру’знее, кукуру’зней, покукуру’зней… смотреть
КУКУРУЗНЫЙ
прил.de (con) maízкукурузное поле — maizal mкукурузная мука — harina de maízкукурузный початок — mazorca f, majorca f
КУКУРУЗНЫЙ
de maïs кукурузные хлопья — flocons m pl de maïs; pop-corn m invarСинонимы: вердепешевый, детской неожиданности, кукурузовый, маисовый, цвет
КУКУРУЗНЫЙ
-ая, -ое.
прил. к кукуруза. Кукурузное поле. Кукурузный стебель.||Приготовленный из кукурузы, с кукурузой.Кукурузные лепешки.Синонимы:
вердепешевый, д… смотреть
Кукурузное поле. Кукурузный стебель.||Приготовленный из кукурузы, с кукурузой.Кукурузные лепешки.Синонимы:
вердепешевый, д… смотреть
КУКУРУЗНЫЙ
корень — КУКУРУЗ; суффикс — Н; окончание — ЫЙ; Основа слова: КУКУРУЗНВычисленный способ образования слова: Суффиксальный∩ — КУКУРУЗ; ∧ — Н; ⏰ — ЫЙ; Сло… смотреть
КУКУРУЗНЫЙ
玉米[的] yùmǐ[de]кукурузная мука — 玉米面кукурузные хлопья — 玉米片Синонимы: вердепешевый, детской неожиданности, кукурузовый, маисовый, цвет
КУКУРУЗНЫЙ
de maïs {mais} кукурузные хлопья — flocons m pl de maïs; pop-corn m invar
КУКУРУЗНЫЙ
Mais- (опр. сл.)кукурузный початок — Maiskolben mСинонимы: вердепешевый, детской неожиданности, кукурузовый, маисовый, цвет
КУКУРУЗНЫЙ
КУКУРУЗНЫЙ прилагательное 1) см. кукуруза , связанный с ним. 2) Свойственный кукурузе , характерный для нее. 3) Предназначенный для уборки, обработки кукурузы . 4) Сделанный, изготовленный из кукурузы …. смотреть
КУКУРУЗНЫЙ
кукурузный Mais. .. кукурузный початок Maiskolben m 1dСинонимы:
вердепешевый, детской неожиданности, кукурузовый, маисовый, цвет
.. кукурузный початок Maiskolben m 1dСинонимы:
вердепешевый, детской неожиданности, кукурузовый, маисовый, цвет
КУКУРУЗНЫЙ
прлde milhoСинонимы: вердепешевый, детской неожиданности, кукурузовый, маисовый, цвет
КУКУРУЗНЫЙ
прил. di granoturco / mais кукурузная каша — (specie di) polenta Итальяно-русский словарь.2003. Синонимы: вердепешевый, детской неожиданности, кукурузовый, маисовый, цвет… смотреть
КУКУРУЗНЫЙ
кукур’узныйСинонимы: вердепешевый, детской неожиданности, кукурузовый, маисовый, цвет
КУКУРУЗНЫЙ
mısır °Синонимы: вердепешевый, детской неожиданности, кукурузовый, маисовый, цвет
КУКУРУЗНЫЙ
кукурузный, ая, -ое кукуруза-га т.; кукурузное поле меке жүгөрү айдалган талаа; кукурузный хлеб загара нан, жүгөрү нан; кукурузный початок жүгөрү сотосу…. смотреть
КУКУРУЗНЫЙ
cornСинонимы: вердепешевый, детской неожиданности, кукурузовый, маисовый, цвет
КУКУРУЗНЫЙ
Кукурузовый кукуру(д)зяний. -ный хлеб — малай (-лая). [Ой малаю, малаю, за хліб тебе не маю (Приказка)]. -ная водка — кукурудзянка.
-ный хлеб — малай (-лая). [Ой малаю, малаю, за хліб тебе не маю (Приказка)]. -ная водка — кукурудзянка.
КУКУРУЗНЫЙ
Начальная форма — Кукурузный, винительный падеж, единственное число, качественное прилагательное, мужской род, неодушевленное
КУКУРУЗНЫЙ
КУКУРУЗНЫЙ кукурузная, кукурузное. Прил. к кукуруза. Кукурузный початок. || Из кукурузы. Кукурузный хлеб.
КУКУРУЗНЫЙ
Кукурузный Кун Кыр Ный Рнк Кук Рык Узы Унр Кныр Кнур Кнр Зырк Зык Зуй Крузый Кыз
КУКУРУЗНЫЙ
-ая, -ое жүгері;- кукурузная мука жүгері ұны;- кукурузный початок жүгері собығы
КУКУРУЗНЫЙ
кукуруз||ныйприл καλαμπο-κίσιος, τοῦ ἀραβοσίτου, τοῦ καλαμπο-κιοῦ.
КУКУРУЗНЫЙ
-ая -ое ж кукуруз …ы; к. початок кукуруз чәкәне
КУКУРУЗНЫЙ ВЕТЕР
—теплый и сухой ветер в степях юго-востока ETC, способствующий созреванию кукурузы. См. Тюркенвипд. Ср. Мельтеми, Траубен-кохер.
КУКУРУЗНЫЙ ЖУК
кукурузный жук
сущ., кол-во синонимов: 1
• жук (372)
Словарь синонимов ASIS. В.Н. Тришин.2013.
.
Синонимы:
жук
В.Н. Тришин.2013.
.
Синонимы:
жук
КУКУРУЗНЫЙ ИНВЕНТАРЬ
машины и орудия, применяемые при выращивании кукурузы с момента ее посадки до момента уборки с поля и обмолота. Сюда входят: кукурузные сеялки, пропашн… смотреть
КУКУРУЗНЫЙ МОТЫЛЕК
просяной червь, конопляный червь, небольшая буровато-желтая или бледно-желтая бабочка. Гусеницы К. м. светло-желтого цвета с темной продольной полоской… смотреть
КУКУРУЗНЫЙ МОТЫЛЕК
кукурузный мотылек сущ., кол-во синонимов: 1 • насекомое (281) Словарь синонимов ASIS.В.Н. Тришин.2013. . Синонимы: насекомое
КУКУРУЗНЫЙ МОТЫЛЕК
Кукурузный мотылек или просяной мотылек (Botys silacealis) — бабочка из семейства Pyralidae (см. Огневки). Самка бледно-желтая, на передних крыльях ржа… смотреть
КУКУРУЗНЫЙ МОТЫЛЕК
или просяной мотылек (Botys silacealis) — бабочка из семейства Pyralidae (см. Огневки). Самка бледно-желтая, на передних крыльях ржаво-бурые поперечные. .. смотреть
.. смотреть
КУКУРУЗНЫЙ ПОЯС
Corn Belt, назв., данное гл. сельскохозяйственному региону США, который простирается через равнины Среднего Запада от штата Огайо до штата Небраска. … смотреть
Генетика маисового полоза
Маисовые полозы являются одними из популярнейших домашних змей. И в первую очередь это оправдано их неприхотливостью и разнообразием окраса. Размер, характер и немыслимое количество морф делает маисового полоза идеальной змеёй не только для начинающих киперов, но и для профессионалов террариумного дела. Но откуда же берётся всё это разноцветное великолепие?
Изначально, природный окрас маисового полоза (чаще всего именуется как “wild type” или “normal”) состоит из комбинации трёх цветов – черного, красного и немного желтого. Эта информация о цвете содержится в ДНК змеи, где гены отвечают за определенные цвета: их качество и количество. И самые простые мутации (морфы) образуются по следующим признакам:
1. Amelanism – полное отсутствие черного пигмента, при котором змеи имеют розовые или красные глаза (потому как из-за отсутствия окраса, просвечиваются кровяные капилляры), а окрас тела варьирует от бело-розового до красного. Чаще всего змей-амеланистов называют “albino”, что не совсем правильно, но более удобно в произношении. Самыми распространенными морфами амеланизма являются – albino (amel), candycane, sunglow, и reverse okeete.
Чаще всего змей-амеланистов называют “albino”, что не совсем правильно, но более удобно в произношении. Самыми распространенными морфами амеланизма являются – albino (amel), candycane, sunglow, и reverse okeete.
2. Hypomelanism – значительное снижение уровня меланина, кайма вокруг сёдел и вентральные чешуйки у таких змей имеют коричневый, серый или светло-коричневый цвет вместо черного. Самые распространенные морфы: hypo и crimson.
3. Anerythrysm – полное отсутствие красного пигмента, змеи обычно светло-серые с тёмными сёдлами, имеют некоторое количество желтого на шее и внизу живота. С помощью этой мутации в комбинации с другими получают морфы snow, ghost и granite.
4. Charcoal – стремление окраса к нейтральным серым и коричневым оттенкам и низкому контрасту рисунка с основным цветом тела, при этом желтый пигмент либо отсутствует вообще, либо присутствует в районе шеи в очень маленьком количестве. Иногда этот тип мутации называют anery B. Основные морфы: pewter, blizzard и phantom.
Иногда этот тип мутации называют anery B. Основные морфы: pewter, blizzard и phantom.
5. Caramel – мутация, подавляющая красный пигмент и заменяющая его на желтый. В сочетании с другими генами образует морфы: butter, amber и gold dust.
6. Lava – мутация, преобразующая черный пигмент. Такие змеи выглядят практически однотонными – рыжие сёдла почти сливаются с основным окрасом змеи, кое-где можно заметить небольшие тёмные вкрапления.
7. Lavender – очень интересная мутация, при которой отсутствует большая часть эритринов и уменьшается количество меланина. Окрас змеи получается вариабельным – от нежно-лавандового до розового с кофейными сёдлами. Наиболее известные морфы: hypo lavender, opal и orchids.
8. Dilute – мутация умеренно преобразующая меланин. В этом случае змеи получаются в нежных постельных тонах. Рисунок как правило нечеткий, состоящий из крупных точек и вкраплений.
на фото dilute caramel
9. Cinder – мутация, удаляющая основной цвет змеи и уменьшающая красный цвет сёдел. Такие змеи выглядят пепельно-серыми или светло-серыми с коричневыми сёдлами. В основном используется в комбинации с другими морфами.
на фото amel cinder
Эти мутации являются рецессивными. И не проявляются в первом поколении. То есть – при скрещивании амеланиста с анеритристом, получится 100% потомства с обыкновенным природным окрасом. Но, если это потомство скрещивать между собой – то можно получить не только детей с обоими признаками, но и другую морфу – snow.
Она образовалась в результате ди-гибридного скрещивания, при котором участвовала не одна пара признаков, а две.
Морфы полученные, в результате ди-гибридного скрещивания: Amber, Amel Cinder (Peppermint), Amel Motley, Amel Stripe, Amel Sunkissed (Inferno), Amel Terrazzo, Anery Caramel, Anery Lavender (Moonstone), Anery Motley, Anery Stripe, Anery Sunkissed, Blizzard, Butter, Caramel Diffused, Caramel Motley, Charcoal Lava (Diamond), Caramel Stripe, Caramel Sunkissed (Honey), Charcoal Motley, Charcoal Stripe, Coral Snow, Charcoal Terrazzo, Diffused Motley, Diffused Stripe, Dilute Anery (Blue), Dilute Motley, Ghost, Golddust, Granite, Hypo Amel, Hypo Cinder, Hypo Diffused, Hypo Lavender, Hypo Motley, Hypo Stripe, Hypo Sunkissed, Ice, Lava Caramel (Topaz), Lava Diffused, Lavamel, Lava Motley, Lavender Diffused, Lavender Motley, Lavender Stripe, Lavender Sunkissed (Orchid), Motley-Stripe, Opal, Pewter, Phantom, Platinum, Snow, Sunkissed Diffused, Topaz, Ultramel.
Так же существует три-гибридное скрещивание, в котором участвуют три гена. В результате этого получаются морфы: Amber Diffused, Amber, Lavender, Amber Motley, Amber Stripe, Blizzard Diffused, Butter Diffused, Butter Motley, Butter Stripe, Caramel Snow, Coral Snow, Dilute Anery Motley, Dilute Anery Stripe, Fire Stripe, Ghost Diffused, Ghost Motley, Ghost Stripe, Golddust Diffused, Golddust Stripe, Granite Motley, Granite Stripe, Hypo Butter, Hypo Lavender Diffused, Hypo Lavender Stripe, Hypo Opal, Hypo Pewter, Ice Diffused, Opal Motley, Opal Stripe, Snow Diffused, Snow Motley, Snow Stripe, Sunkissed Snow, Ultra Caramel Motley, Ultramel Anery, Ultramel Diffused, Ultramel Lavender.
Для примера рассмотрим морфу Caramel Snow. Мы уже знаем, что caramel – это простая рециссивная мутация, а snow – это морфа, полученная в результате ди-гибридного скрещивания anery и amel. Получается, что Caramel Snow – это комбинация трёх признаков – anery, amel и caramel. Всё просто.
Всё просто.
Так же в названиях морф часто встречаются слова: motley и stripe. Что же это такое? Если воспользоваться переводчиком, то будет понятно, что motley – это пятнистый, а stripe – это полоса. Таким образом – эти два слова обозначают тип узора. Stripe – превращает привычную форму сёдел в продольные полосы, которые непрерывно идут от головы до хвоста, либо прерываются, или вовсе исчезают в середине. А motley расширяет края сёдел, заключая вовнутрь основной цвет змеи, в таком случае образуются пятна, имеющие различную форму от круглых, до вытянутых в толстую полосу.
Эти признаки также являются рецессивными.
Маисовый полоз: уход и содержание, преимущества и недостатки
На сегодняшний день среди владельцев террариумов набирает популярность новая рептилия — маисовый полоз. Что же это за зверь такой и с чем его едят?
Есть его, конечно же, не стоит — а вот разобраться, что это за рептилия такая, как правильно его содержать и ухаживать, пожалуй, нужно.
Как выглядит маисовый полоз?
Малыши рептилии имеют длину от 20 до 40 сантиметров, а взрослая особь достигает размеров 150 сантиметров. Как уже упоминалось, отличительная черта змеи — уникальный окрас. Орнаменты полоза очень разнообразны, а цветовая гамма, на любой «вкус и цвет» — чёрный, красный, оранжевый, кремовый, малиновый, розовый, коралловый, вишнёвый, коричневый, белый…
Где ещё можно найти змею с таким разнообразием окрасов? Окити — представители классической окраски ярких, насыщенных цветов. Есть и представители, не имеющие в своём окрасе ярких цветов — амеланисты. Это белые и кремовые змеи с небольшими оттенками жёлтого или оранжевых цветов. Полоз альбинос имеет светло-кремовый цвет с еле заметными тёмными полосками и выделяющиеся глаза красного цвета.
Почему же эта рептилия стала столь популярна среди террариумистов?
- Они абсолютно не агрессивны.
- Необычный и разнообразный окрас.

- Небольшой размер в сравнении с другими рептилиями.
- Очень быстро привыкают «к рукам».
- Полосы не кусаются.
- Живут от 10 до 20 лет, в зависимости от условий содержания.
- Лёгкие и неприхотливые в содержании.

Есть у маисового полоза и несколько недостатков:
- Подвержен поражению бактериями, поэтому очень важно следить за чистотой террариума и состоянием здоровья рептилии.
- Очень юркие и любят осваивать новое пространство — поэтому способны убежать. Дабы избежать этого нужно заранее обустроить жилище для будущего питомца.
Образ жизни маисового полоза
Взрослые особи хорошо ползают и могут залезть на довольно большую высоту, а вот у молоденьких сил на это не хватает. Зимой эти рептилии впадают в спячку и просыпаются лишь с приходом весенних тёплых деньков. Начинают проявлять активность примерно в середине мая, когда трава уже довольно высокая. В естественной среде обитания на зимовку они заползают в укромные места: под брёвна, в расщелины скал и тому подобные.
В террариуме также будут искать укромный уголок, где полоза никто не побеспокоит. Наивысшая точка активности у полозов наступает ночью или в предрассветное время, днём же они предпочитают отдыхать. Излюбленное занятие рептилии — охота на грызунов, ящериц, небольших птиц. Настигнув цель, полоз крепко сжимает её и душит.
Маисовый полоз содержание
Для содержания маисового полоза вполне хватит небольшого террариума (около 80 литров, для детёнышей хватит 40–50 литров), в котором необходимо поддерживать температуру около 25–29 градусов и влажность порядка 60% (в период линьки — 80%). И также террариум должен хорошо вентилироваться.
Для обогрева «домика» можно использовать специальные термоковрики, термошнуры, лампы накаливания и зеркальные лампы, а для поддержания нужного уровня влажности — кювету с увлажнённым сфагнумом. Если для обогрева вы выбрали термошнур, то прокладывать его лучше под дном террариума, чем в самом грунте. А если ваш выбор пал на лампу — её нужно установить так, чтобы рептилия не обожглась, желательно отгородить её стеклом или сеткой. На ночь обогрев террариума можно отключать — если температура в нём не опускается ниже 22 градусов. И не забудьте установить в террариуме несколько градусников для контроля над температурой, так как температура очень важна для нормальной жизнедеятельности и пищеварения вашего питомца.
На ночь обогрев террариума можно отключать — если температура в нём не опускается ниже 22 градусов. И не забудьте установить в террариуме несколько градусников для контроля над температурой, так как температура очень важна для нормальной жизнедеятельности и пищеварения вашего питомца.
На дно «жилища» нужно насыпать крупный песок, бумагу, дроблёную кору, гравий или же специальный кокосовый субстрат. Тут есть один нюанс — если вы выбрали для покрытия опилки, мелкую стружку или кокосовый субстрат, то кормить рептилию придётся в другом месте из-за угрозы попадания в его организм частичек грунта вместе с пищей.
В обязательном порядке нужно обустроить рептилии укромный уголок — домик, декоративное бревно, полочка…выбор огромен. Только помните, что размер укрытий должен позволять полозу полностью в них спрятаться. В идеале они должны иметь несколько отверстий, дабы змее было комфортно вползать/выползать. Будет неплохим развлечением для полоза и преодолевания всяко-разных веточек, палочек, коряг. Освещать террариум необходимо лампами дневного света.
Освещать террариум необходимо лампами дневного света.
Маисовые полозы, как и многие другие рептилии во время линьки, стремятся в воду — полежать, отмокнуть, поплавать. Так что не забудьте поставить красавцу своеобразную «ванну». Ну и, конечно же, не забываем про устойчивую поилку с высокими стенами.
Можно ли содержать несколько маисовых полозов в одном террариуме?
Конечно же, можно, и даже нужно, если размеры вашего террариума это позволяют! На одного самца в террариуме допускаются максимум две самки. Внимание! Когда полозы выходят из зимней спячке их необходимо временно рассадить. И также кормить придётся всех по отдельности.
Рацион маисового полоза
Немаловажный аспект в уходе за маисовым полозом составляет его питание.
Кормить полоза нужно один раз в пять дней — после того, как съеденное ранее переварилось и вышло наружу. Взрослые особи в среднем едят 1 раз в неделю. Помните, что у змей отсутствует чувство меры, поэтому не перекармливайте!
Основной рацион рептилии составляют мыши — количество зависит от размера и возраста змеи (1–2 крупные мыши). Можно докармливать полоза специальным размороженным кормом. Не стоит допускать, чтобы рептилия сама ловила себе мышку, так как есть вероятность инфекции и отравления. Взрослым особям желательно иногда разнообразить рацион некрупными цыплятами, перепелятами, песчанками или хомячками, но часто этого делать не нужно.
Можно докармливать полоза специальным размороженным кормом. Не стоит допускать, чтобы рептилия сама ловила себе мышку, так как есть вероятность инфекции и отравления. Взрослым особям желательно иногда разнообразить рацион некрупными цыплятами, перепелятами, песчанками или хомячками, но часто этого делать не нужно.
Но также не стоит забывать про различные минеральные добавки — кальций, яичная скорлупа. А вот концентрированные витамины стоит давать нечасто — одного раза в месяц будет предостаточно. Не стоит экспериментировать и с мясом типа свинины или говядины, и рыбы — пищеварительная система маисовых полозов не приспособлена к такой пище, соответственно питомец от неё может заболеть и даже умереть. Всегда проверяйте наличие воды в поилке питомца и каждый день меняйте её, это является одним из важнейших условий содержания маисового полоза! В воду иногда можно подмешивать минеральную воду типа «Боржоми», как дополнительный источник витаминов и минералов.
Кормление малыша полоза не сильно отличается от взрослой особи — тут важно не допускать, чтобы детёныш голодал. Давать корм нужно один раз в 4–5 дней, опять же при условии полного опорожнения от предыдущей порции. Совсем маленьких полозов, до шести месяцев кормят только новорождёнными мышатами.
После кормления маисового полоза тревожить не рекомендуется, так как рептилия может срыгнуть. Если такое всё же произошло нужно сделать перерыв в кормлении не менее недели.
В период линьки маисового полоза кормить не нужно.
Не стоит протягивать еду для рептилии голыми руками, т. к. маисовый полоз заглатывает пищу целиком и вполне может прихватить и руку.
Возможно ли разведение маисовых полозов в домашних условиях?
Как уже упоминалось ранее, разведение этих змей дома не только возможно, но и зачастую более эффективно, чем в условиях дикой природы.
Когда же эти рептилии способны к размножению? Половое созревание у них наступает в возрасте двух-трёх лет. После достижения половозрелового возраста нужно дождаться, когда рептилии выйдут из зимовки, и в течение двух — трёх недель в корм добавлять витамин «Е». Затем подсадить друг к другу.
После достижения половозрелового возраста нужно дождаться, когда рептилии выйдут из зимовки, и в течение двух — трёх недель в корм добавлять витамин «Е». Затем подсадить друг к другу.
Спаривание происходит в течение 10–20 минут, после оного самки откладывают яйца через 1–2 месяца. В террариуме необходимо установить кювету со сфагнумом — там самка сделает кладку. Кладка обычно состоит из 6–35 яиц.
Яйца необходимо поместить в инкубатор, где нужно поддерживать температуру 27–29 градусов. Через 50–75 дней на свет появятся малыши маисовых полозов. Питаться детёныши начинают после первой линьки.
При содержании в домашних условиях можно добиться повторной кладки — для этого необходимо дополнительно стимулировать самцов и самок. В дикой же среде обитания такого явления не наблюдалось.
Вот мы и рассмотрели принципы содержания и ухода маисового полоза. Исходя из всего вышеперечисленного становится понятным, почему эта рептилия так полюбилась заводчикам змей — она неприхотлива в еде и уходе и не доставляет особых хлопот. Для новичков, которые никогда ранее не имели дела со змеями, но хотят заняться этим интересным делом — это идеальная змея. Неприхотлив, миролюбив — что ещё нужно? Конечно же, как и в случае с другими рептилиями и животными — приобретать лучше детёныша, так как его будет легче приручить. Детёныши достаточно быстро начинают признавать хозяина и любят сопровождать его во всех домашних делах.
Для новичков, которые никогда ранее не имели дела со змеями, но хотят заняться этим интересным делом — это идеальная змея. Неприхотлив, миролюбив — что ещё нужно? Конечно же, как и в случае с другими рептилиями и животными — приобретать лучше детёныша, так как его будет легче приручить. Детёныши достаточно быстро начинают признавать хозяина и любят сопровождать его во всех домашних делах.
Post Views: 1 083
границ | Транскриптомный анализ генов времени цветения в условиях засушливого стресса в листьях кукурузы
Введение
Кукуруза ( Zea mays L.) — основная сельскохозяйственная культура, используемая во всем мире в качестве источника пищи, топлива и корма для животных. Кукуруза служит ключевым ресурсом для экономического и промышленного применения, в дополнение к ее роли в качестве пищи. Время цветения является важной характеристикой производства кукурузы, которая, как известно, влияет на продуктивность и качество семян. Что касается воспроизводства кукурузы, от 1 недели до шелушения и до 2 недель после шелушения представляют собой ключевой период, в течение которого определяются аборты семязачатков и скорость оплодотворения, а также появляются зерна и колосья.Пути, необходимые для цветения, имеют системы реагирования, которые опосредуют выживание при различных стрессах. В ответ на стресс, такой как засуха, пути цветения ускоряются, чтобы быстрее производить цветы и семена (Franks et al., 2007; Bernal et al., 2011; Franks, 2011; Krasensky and Jonak, 2012). Стресс от засухи имеет много негативных последствий, таких как снижение доступности углерода, опадение цветков, гибель пыльцы, аборты яйцеклеток и интервал цветения-шелушения (ASI) во время репродуктивных стадий (Hall et al., 1984; Блюм, 1996; Андраде и др., 1999). Увеличение ASI, вызванное засухой, негативно влияет на норму внесения удобрений, наполнение зерен, качество и вес семян (Byrne et al., 1995; Bolaños and Edmeades, 1996; Edmeades et al., 1997; Bruce et al.
Что касается воспроизводства кукурузы, от 1 недели до шелушения и до 2 недель после шелушения представляют собой ключевой период, в течение которого определяются аборты семязачатков и скорость оплодотворения, а также появляются зерна и колосья.Пути, необходимые для цветения, имеют системы реагирования, которые опосредуют выживание при различных стрессах. В ответ на стресс, такой как засуха, пути цветения ускоряются, чтобы быстрее производить цветы и семена (Franks et al., 2007; Bernal et al., 2011; Franks, 2011; Krasensky and Jonak, 2012). Стресс от засухи имеет много негативных последствий, таких как снижение доступности углерода, опадение цветков, гибель пыльцы, аборты яйцеклеток и интервал цветения-шелушения (ASI) во время репродуктивных стадий (Hall et al., 1984; Блюм, 1996; Андраде и др., 1999). Увеличение ASI, вызванное засухой, негативно влияет на норму внесения удобрений, наполнение зерен, качество и вес семян (Byrne et al., 1995; Bolaños and Edmeades, 1996; Edmeades et al., 1997; Bruce et al. , 2002). По этой причине ASI часто используется в качестве индекса засухоустойчивости.
, 2002). По этой причине ASI часто используется в качестве индекса засухоустойчивости.
Для улучшения продуктивности в условиях засушливого стресса было предпринято несколько попыток улучшить сорта с использованием рекомбинантных инбредных линий (RIL), молекулярных маркеров и картирования локусов количественных признаков (QTL) (Cattivelli et al., 2008; Ю и др., 2008; Макмаллен и др., 2009). Однако сложность генома кукурузы и отсутствие знаний о конкретных генетических механизмах, лежащих в основе устойчивости к засухе, остаются серьезной проблемой. В соответствии с разнообразием этих сильно гетерозиготных растений и их геномной сложностью было обнаружено, что разные генотипы демонстрируют разные системы реагирования на стресс от засухи (Shinozaki and Yamaguchi-Shinozaki, 2007; Hansey et al., 2012; Jiang et al., 2012). . Поэтому, чтобы улучшить наше понимание взаимосвязи между засухой и урожайностью, наша цель состояла в том, чтобы охарактеризовать механизмы, связанные с реакцией на засуху, которые влияют на время цветения.
Время цветения, определяющее ASI кукурузы, тесно связано с генами времени цветения. Известно, что такие гены, как FLOWERING LOCUS C (FLC), CONSTANS (CO), GIGANTEA (GI) и SUPPRESSOR of OVEREXPRESSION of CONSTANS1 (SOC1), регулируют время цветения (Kim et al., 2006; Buckler et al., 2009; Jung and Müller, 2009; Kong et al., 2010; Endo-Higashi and Izawa, 2011; Hung et al., 2012; Ito et al., 2012; Li et al., 2016). Экспрессия этих генов контролируется фотопериодом, светочувствительностью, циркадными ритмами и различными стрессами (Johnson et al., 1994б; Датта, 2006 г.; Форнара и др., 2009 г.; Юнг и Мюллер, 2009 г.; Мэн и др., 2011; Хунг и др., 2012; Коэльо и др., 2013; Чао и др., 2014). Точно так же известно, что засуха вносит изменения в ASI, поэтому засуха может влиять на гены времени цветения. Наблюдались изменения уровня экспрессии генов времени цветения из-за засухи (Franks et al., 2007; Zhuang et al., 2007; Kakumanu et al., 2012; Corrales et al., 2014; Kooyers, 2015). Более подробный анализ паттерна экспрессии генов может привести к улучшению нашего понимания регуляции процессов, связанных со временем цветения в условиях засухи.
Исследования экспрессии генов, которые характеризуют реакцию кукурузы на стресс от засухи, были предприняты в различных тканях (Zheng et al., 2004; Poroyko et al., 2007; Zhuang et al., 2007) и на нескольких стадиях (Zheng et al. , 2004; Юэ и др., 2008; Гумберт и др., 2013). С появлением технологий секвенирования РНК (RNA-seq) и публикацией эталонного генома (Schnable et al., 2009) у нас теперь есть доступ к большому количеству информации о разнообразии генома и экспрессии генов у кукурузы, включая структурные вариации, различные системы реагирования и альтернативные события сплайсинга (Wang et al., 2009; Джайн, 2011). Аннотации генов и последовательности, включенные в эталонный геном, сделали возможным улучшение геномных исследований. Недавно секвенирование РНК использовалось для характеристики транскрипционных изменений, вызванных абиотическим стрессом (Krasensky and Jonak, 2012; Frey et al., 2015; Liu et al., 2015), и было обнаружено, что идентифицированные дифференциально экспрессируемые гены (DEG) быть полезным для прогнозирования различий в толерантности между сортами кукурузы. Альтернативный сплайсинг (АС) также играет важную роль в реакциях на стресс, и, по оценкам, события АС происходят для более чем 60% интронсодержащих генов в Arabidopsis thaliana (Филичкин и др., 2010). Также известно, что АС по-разному возникает в разных тканях, на разных стадиях развития и в ответ на стресс у растений (Li et al., 2013; Staiger and Brown, 2013; Garg et al., 2016; Shankar et al., 2016). .
Альтернативный сплайсинг (АС) также играет важную роль в реакциях на стресс, и, по оценкам, события АС происходят для более чем 60% интронсодержащих генов в Arabidopsis thaliana (Филичкин и др., 2010). Также известно, что АС по-разному возникает в разных тканях, на разных стадиях развития и в ответ на стресс у растений (Li et al., 2013; Staiger and Brown, 2013; Garg et al., 2016; Shankar et al., 2016). .
В этом исследовании мы использовали секвенирование РНК для выявления вариаций транскрипции между растениями кукурузы, подвергавшимися воздействию обильного полива (WW) и стресса от засухи (DS) в условиях короткого дня. Полученные данные секвенирования РНК были проанализированы с использованием инструментов биоинформатики для выявления изменений в экспрессии генов и альтернативного сплайсинга в условиях засушливого стресса.Гены с дифференциальной экспрессией подвергали анализу BLAST для прогнозирования их функций. Затем мы сосредоточились на генах, реагирующих на засуху, которые, как известно или предполагается, участвуют во времени цветения, и проанализировали потенциал этих генов по изменению времени цветения в ответ на стресс, вызванный засухой.
Материалы и методы
Рост растений и условия засухи
растения кукурузы ( Zea mays сорта B73) выращивали в 10-литровых горшках (37 × 37,5 × 22 см). Растения содержали в теплице и выращивали в условиях короткого дня до тех пор, пока верхушка метелки не стала видна.Условия короткого дня устанавливались при 12-часовом светлом и 12-часовом темном и умеренном климате от 26 до 28°С днем и 23-25°С ночью. Когда метелка была видна, растения делили на две группы: хорошо поливаемые (WW) и засухоустойчивые (DS). Горшки WW поддерживали при потенциалах >-0,2 МПа (содержание влаги в почве 15–20 %) с орошением, тогда как горшки DS поддерживали при потенциалах <-1,5 МПа (содержание влаги в почве 5–8 %). Когда началось опадение пыльцы, ткань листа собирали с верхнего листа с воротничком (флагового листа) с трех разных растений в каждой группе повторов и условиях.В результате из-за засухи изменения ASI составили 3–4 дня, а образцы из каждого условия были собраны в разные дни. В среднем группа ДС находилась более 15 дней в условиях засухи. Все листья были заморожены в жидком азоте сразу после сбора, а затем перенесены в морозильную камеру для хранения при температуре -80°C.
В среднем группа ДС находилась более 15 дней в условиях засухи. Все листья были заморожены в жидком азоте сразу после сбора, а затем перенесены в морозильную камеру для хранения при температуре -80°C.
Выделение РНК и секвенирование РНК
Для создания РНК-библиотек листья, собранные с растений WW и DS, подвергали выделению РНК с биологической репликацией.Тотальную РНК получали из каждого листа с использованием набора RNeasy Plant Mini Kit (Qiagen). Качество и целостность РНК оценивали с использованием РНК-наночипа 2100 Bioanalyzer (Agilent Technologies) перед секвенированием РНК. Затем выполняли секвенирование РНК с использованием платформы Illumina HiSeq.
Биоинформатика
Все чтения из каждого образца были сопоставлены с эталонным геномом кукурузы [сборка аннотации B73_RefGen_v3 (5b+)] с использованием параметров по умолчанию в Tophat2. Методология Tophat облегчает идентификацию событий сплайсинга.Выровненные последовательности из Tophat2 (Kim et al. , 2013) были собраны отдельно с использованием Cufflinks 2.2.1 (Trapnell et al., 2012). Cufflinks собирает определенные изоформы на основе событий альтернативного сплайсинга. Результаты Cufflinks сравнивали с аннотацией эталонного генома (http://plants.ensembl.org/Zea_mays/) с использованием Cuffmerge. Коды классов, полученные из выходных данных Cuffmerge, использовались для идентификации новых изоформ, возникающих в результате недавно идентифицированных соединений сплайсинга и межгенных транскриптов.
, 2013) были собраны отдельно с использованием Cufflinks 2.2.1 (Trapnell et al., 2012). Cufflinks собирает определенные изоформы на основе событий альтернативного сплайсинга. Результаты Cufflinks сравнивали с аннотацией эталонного генома (http://plants.ensembl.org/Zea_mays/) с использованием Cuffmerge. Коды классов, полученные из выходных данных Cuffmerge, использовались для идентификации новых изоформ, возникающих в результате недавно идентифицированных соединений сплайсинга и межгенных транскриптов.
Дифференциально экспрессируемые гены (DEG) были идентифицированы путем подсчета значений фрагментов на тысячу оснований транскрипта на миллион картированных считываний (FPKM).DEG были определены как гены, имеющие коэффициент ложных открытий (FDR) (Benjamini and Yekutieli, 2001) <0,001 и абсолютное значение логарифмического 2 кратного изменения >1. Чтобы дополнительно охарактеризовать гены, идентифицированные как дифференциально экспрессирующиеся в ответ на стресс от засухи, DEG и консенсусные последовательности изоформ были сопоставлены с классификациями GO с использованием Blast2GO (Conesa and Götz, 2008). Были проанализированы три категории аннотаций GO: биологический процесс, молекулярная функция и клеточный компонент. Анализ обогащения GO был выполнен с помощью плагинов BiNGO для Cytoscape (Maere et al., 2005) с использованием гипергеометрического критерия и метода коррекции Бонферрони. Поправку Бонферрони использовали для коррекции значения P с отсечкой 0,05. Для выявления альтернативных событий сплайсинга, связанных со стрессом от засухи, данные Cuffdiff были проанализированы с использованием модуля spliceR в R (Vitting-Seerup et al., 2014).
Были проанализированы три категории аннотаций GO: биологический процесс, молекулярная функция и клеточный компонент. Анализ обогащения GO был выполнен с помощью плагинов BiNGO для Cytoscape (Maere et al., 2005) с использованием гипергеометрического критерия и метода коррекции Бонферрони. Поправку Бонферрони использовали для коррекции значения P с отсечкой 0,05. Для выявления альтернативных событий сплайсинга, связанных со стрессом от засухи, данные Cuffdiff были проанализированы с использованием модуля spliceR в R (Vitting-Seerup et al., 2014).
Количественная полимеразная цепная реакция в реальном времени (qRT-PCR) Валидация данных секвенирования РНК
РНК, выделенных из групп WW и DS, использовали для создания библиотеки кДНК.Синтез первой цепи осуществляли с использованием набора для синтеза кДНК PrimeScript First Strand (Takara) и случайных праймеров в соответствии с процедурами, рекомендованными производителем. qRT-PCR проводили с использованием системы обнаружения ПЦР в реальном времени CFX Connect™ (Bio-Rad). Метод CT использовали для количественной оценки изменений в экспрессии генов, а рРНК 18s кукурузы и Actin1 служили эталонами для нормализации. Экспрессию анализировали для каждого из выбранных генов в обоих условиях (WW и DS). Три независимых биологических повтора каждого образца подвергали анализу qRT-PCR.Чтобы определить относительные кратные изменения, экспрессию генов нормализовали до значений CT для эталонных генов. Значения CT для каждого гена в каждом состоянии рассчитывали с использованием метода ΔΔCT, как описано ранее (Livak and Schmittgen, 2001).
Метод CT использовали для количественной оценки изменений в экспрессии генов, а рРНК 18s кукурузы и Actin1 служили эталонами для нормализации. Экспрессию анализировали для каждого из выбранных генов в обоих условиях (WW и DS). Три независимых биологических повтора каждого образца подвергали анализу qRT-PCR.Чтобы определить относительные кратные изменения, экспрессию генов нормализовали до значений CT для эталонных генов. Значения CT для каждого гена в каждом состоянии рассчитывали с использованием метода ΔΔCT, как описано ранее (Livak and Schmittgen, 2001).
Результаты
Анализ секвенирования РНК выявил изменения в транскриптоме кукурузы в ответ на стресс, вызванный засухой
образца РНК из хорошо политых (WW) и подверженных засухе (DS) растений подвергали секвенированию РНК с использованием прибора Illumina Hi-Seq 2000.Всего было проанализировано шесть образцов, включая три биологические повторности для каждого условия. Для каждой повторности было получено 99–113 млн прочтений. Для сравнительного анализа было использовано более 280 миллионов парных прочтений, примерно по 47 миллионов прочтений для каждого образца (таблица 1). Из этих прочтений 86,7–89,0% могут быть сопоставлены с эталонным геномом ( Zea mays AGPv3.30) как уникальные и множественные совпадения. Сопоставленные чтения были проанализированы с использованием пайплайна Cufflinks (Trapnell et al., 2012) для выявления DEG, событий альтернативного сплайсинга и новых транскриптов. Значения экспрессии, связанные с генами и транскриптами, рассчитывали с использованием модели фрагментов на тысячу оснований экзона на миллион картированных фрагментов (FPKM). Также экспрессию ZmDREB2A использовали для проверки растений, испытывающих стресс от засухи (рис. S1).
Для сравнительного анализа было использовано более 280 миллионов парных прочтений, примерно по 47 миллионов прочтений для каждого образца (таблица 1). Из этих прочтений 86,7–89,0% могут быть сопоставлены с эталонным геномом ( Zea mays AGPv3.30) как уникальные и множественные совпадения. Сопоставленные чтения были проанализированы с использованием пайплайна Cufflinks (Trapnell et al., 2012) для выявления DEG, событий альтернативного сплайсинга и новых транскриптов. Значения экспрессии, связанные с генами и транскриптами, рассчитывали с использованием модели фрагментов на тысячу оснований экзона на миллион картированных фрагментов (FPKM). Также экспрессию ZmDREB2A использовали для проверки растений, испытывающих стресс от засухи (рис. S1).
Таблица 1. Сводка данных секвенирования РНК из образцов WW и DS .
Транскриптомный анализ генов, реагирующих на засуху
Чтобы охарактеризовать вариации транскрипции, возникающие в ответ на стресс от засухи, мы провели дифференциальный анализ экспрессии генов. В общей сложности 34 002 гена были экспрессированы как в условиях WW, так и в условиях DS. Из этих генов 954 прошли тест Каффдиффа и были признаны генами, реагирующими на засуху. DEG, которые демонстрировали log 2 -кратное изменение >1, включали 617 генов (рис. 1), 358 из которых были активированы, а 259 из которых были подавлены (рис. 1, таблица S1). Из DEG 512 можно было сопоставить с эталонным геномом, а 105 — нет.
В общей сложности 34 002 гена были экспрессированы как в условиях WW, так и в условиях DS. Из этих генов 954 прошли тест Каффдиффа и были признаны генами, реагирующими на засуху. DEG, которые демонстрировали log 2 -кратное изменение >1, включали 617 генов (рис. 1), 358 из которых были активированы, а 259 из которых были подавлены (рис. 1, таблица S1). Из DEG 512 можно было сопоставить с эталонным геномом, а 105 — нет.
Рисунок 1. Сводная информация о дифференциально экспрессируемых генах, событиях альтернативного сплайсинга и новых транскриптах (NT) в условиях засухи . (A) Количество генов повышающей регуляции, (B) количество генов понижающей регуляции, (C) количество событий альтернативного сплайсинга, классифицированных по каждому типу.
Всего было идентифицировано 92 438 альтернативно сплайсированных транскриптов. SpliceR использовали для идентификации значительно измененных изоформ. Фильтрация привела к идентификации 50 354 общих событий AS, 67 событий AS, специфичных для WW, и 82 событий AS, специфичных для DS (рис. 1C). Альтернативные сайты терминации транскрипции (ATTS) и альтернативные сайты начала транскрипции (ATSS) были наиболее преобладающими событиями AS, составляя 11 943 и 9 465 событий соответственно (рис. 1C).Другие идентифицированные события AS включали 9327 событий альтернативного 3′-сайта сплайсинга (A3), 6185 событий альтернативного 5′-сайта сплайсинга (A5), 8982 события пропуска/удержания интрона (ISI) и 3786 событий пропуска/включения экзона (ESI). Типы событий AS варьировались в зависимости от условий, при этом типы A3, A5 и ESI преобладали в условиях засухи. Всего было идентифицировано 177 чувствительных к засухе изоформ, имеющих q < 0,05. Чтобы охарактеризовать, были ли альтернативно сплайсированные транскрипты значительно дифференциально экспрессированы в условиях засухи, мы выбрали 126 транскриптов с log 2 кратными изменениями >1 (таблица S2).
Фильтрация привела к идентификации 50 354 общих событий AS, 67 событий AS, специфичных для WW, и 82 событий AS, специфичных для DS (рис. 1C). Альтернативные сайты терминации транскрипции (ATTS) и альтернативные сайты начала транскрипции (ATSS) были наиболее преобладающими событиями AS, составляя 11 943 и 9 465 событий соответственно (рис. 1C).Другие идентифицированные события AS включали 9327 событий альтернативного 3′-сайта сплайсинга (A3), 6185 событий альтернативного 5′-сайта сплайсинга (A5), 8982 события пропуска/удержания интрона (ISI) и 3786 событий пропуска/включения экзона (ESI). Типы событий AS варьировались в зависимости от условий, при этом типы A3, A5 и ESI преобладали в условиях засухи. Всего было идентифицировано 177 чувствительных к засухе изоформ, имеющих q < 0,05. Чтобы охарактеризовать, были ли альтернативно сплайсированные транскрипты значительно дифференциально экспрессированы в условиях засухи, мы выбрали 126 транскриптов с log 2 кратными изменениями >1 (таблица S2). Из этих транскриптов AS 82 (из 76 генов) были активированы, а 44 (из 43 генов) подавлены (рис. 1). Как правило, гены с изоформами AS также идентифицировали как DEG; однако было обнаружено, что 14 генов подвергаются только дифференциальному сплайсингу. Большинство генов имели одну изоформу с дифференциальной экспрессией, но шесть генов с повышенной экспрессией и один ген с подавленной экспрессией имели две изоформы с дифференциальной экспрессией. Было обнаружено, что некоторые гены имеют новые транскрипты, которые по-разному регулируются в условиях засухи.Из транскриптов с повышенной экспрессией 14 были новыми транскриптами, в том числе девять транскриптов, связанных с DEG (рис. 1А). Напротив, шесть новых транскриптов были подавлены (рис. 1В).
Из этих транскриптов AS 82 (из 76 генов) были активированы, а 44 (из 43 генов) подавлены (рис. 1). Как правило, гены с изоформами AS также идентифицировали как DEG; однако было обнаружено, что 14 генов подвергаются только дифференциальному сплайсингу. Большинство генов имели одну изоформу с дифференциальной экспрессией, но шесть генов с повышенной экспрессией и один ген с подавленной экспрессией имели две изоформы с дифференциальной экспрессией. Было обнаружено, что некоторые гены имеют новые транскрипты, которые по-разному регулируются в условиях засухи.Из транскриптов с повышенной экспрессией 14 были новыми транскриптами, в том числе девять транскриптов, связанных с DEG (рис. 1А). Напротив, шесть новых транскриптов были подавлены (рис. 1В).
GO Обогащение дифференциально экспрессируемых генов и транскриптов
Консенсусные последовательности для генов, реагирующих на засуху, и транскрипты, полученные в результате секвенирования РНК, были подвергнуты Blast2Go для дальнейшего анализа функции генов. Многие из DEG, которые мы идентифицировали, были известными генами, чувствительными к стрессу.Почти все идентифицированные нами чувствительные к засухе гены экспрессировались на уровнях, сходных с предыдущими отчетами, характеризующими экспрессию генов в ответ на засушливый стресс с использованием полногеномной ассоциации и картирования QTL (Shinozaki and Yamaguchi-Shinozaki, 2007; Yue et al., 2008; Setter et al., 2010; Kakumanu et al., 2012; Thirunavukkarasu et al., 2014). Термины GO, связанные с биологическим процессом для DEG, включали реакцию на стресс, реакцию на эндогенный стимул, фотосинтез, хроматическое связывание и реакцию на внешний стимул, которые соответствуют стрессу от засухи (рис. 2А).Термины GO, относящиеся к репродуктивным стадиям, включали развитие цветка (шесть генов), репродукцию (четыре гена), взаимодействие пыльцы и пестика (один ген) и постэмбриональное развитие (один ген) (таблица S3). Термины GO из категории молекулярных функций, связанных с реакцией на стресс, включают связывание углеводов, транспортную активность и связывание хроматина.
Многие из DEG, которые мы идентифицировали, были известными генами, чувствительными к стрессу.Почти все идентифицированные нами чувствительные к засухе гены экспрессировались на уровнях, сходных с предыдущими отчетами, характеризующими экспрессию генов в ответ на засушливый стресс с использованием полногеномной ассоциации и картирования QTL (Shinozaki and Yamaguchi-Shinozaki, 2007; Yue et al., 2008; Setter et al., 2010; Kakumanu et al., 2012; Thirunavukkarasu et al., 2014). Термины GO, связанные с биологическим процессом для DEG, включали реакцию на стресс, реакцию на эндогенный стимул, фотосинтез, хроматическое связывание и реакцию на внешний стимул, которые соответствуют стрессу от засухи (рис. 2А).Термины GO, относящиеся к репродуктивным стадиям, включали развитие цветка (шесть генов), репродукцию (четыре гена), взаимодействие пыльцы и пестика (один ген) и постэмбриональное развитие (один ген) (таблица S3). Термины GO из категории молекулярных функций, связанных с реакцией на стресс, включают связывание углеводов, транспортную активность и связывание хроматина.
Рисунок 2. Анализ обогащения чувствительных генов и транскриптов с использованием терминов GO из GO Slim . Значительно перепредставленные термины GO Slim были визуализированы BiNGO как приложение в Cytoscape.Размер узла был пропорционален количеству целей в категории GO. Цвет узла представляет значимость обогащения: более высокая значимость была представлена более глубоким цветом. (A) Результат DEG (B) Результат ответных транскриптов.
Известные гены, реагирующие на засуху, обнаруженные в наших результатах, включали гены, кодирующие альфа- и бета-амилазы (GRMZM2G138464 и GRMZM2G450125), которые расщепляют крахмал — процесс, который, как было показано, активируется при стрессе от засухи (Krasensky and Jonak, 2012; Праш и др., 2015). Два гена, вовлеченные в путь биосинтеза АБК (GRMZM5G858784 и GRMZM2G179147), демонстрируют повышенный уровень транскриптов в условиях засухи (Yue et al., 2008; Urano et al., 2009). Мы также обнаружили, что экспрессия гена, кодирующего сахарозосинтазу 7 (GRMZM2G060583), подавлялась в условиях засухи (таблица S1). В предыдущем исследовании гены, кодирующие синтазу сахарозы, подавлялись в ткани завязи, но не проявляли опосредованных засухой изменений в меристеме листьев (Kakumanu et al., 2012).
Мы также обнаружили, что экспрессия гена, кодирующего сахарозосинтазу 7 (GRMZM2G060583), подавлялась в условиях засухи (таблица S1). В предыдущем исследовании гены, кодирующие синтазу сахарозы, подавлялись в ткани завязи, но не проявляли опосредованных засухой изменений в меристеме листьев (Kakumanu et al., 2012).
В дополнение к DEG мы обнаружили 126 транскриптов со значительно отличающейся экспрессией в ответ на засуху. Подобно DEGs, транскрипты, которые, как известно, участвуют в реакции на засуху, обнаруживают измененную регуляцию в условиях засухи. Например, бета-амилаза (GRMZM2G450125), хитиназа (GRMZM2G005633), каротиноидгидроксилаза (GRMZM2G164318), транскрипты WRKY (GRMZM2G120320 и GRMZM2G018487) и белки теплового шока (GRMZM2G024668 и GRMZM2G428391) активировались засухой.Сообщалось, что эти белки играют важную роль в реакции на засушливый стресс (Hu et al., 2009; Banerjee and Roychoudhury, 2015; Prasch et al., 2015). Дифференциально экспрессируемые изоформы были связаны со многими из тех же терминов GO, что и DEG. Термины GO биологического процесса, связанные с дифференциально экспрессируемыми изоформами, включают ответ на стресс, ответ на абиотический стимул и ответ на эндогенный стимул. Мы также идентифицировали несколько новых изоформ транскриптов, индуцированных стрессом. Из 14 новых изоформ, выставляемых значительно другое выражение, изоформы шести транскриптов (GRMZM2G162598_T01, GRMZM2G045239_T01, GRMZM2G163809_T02, GRMZM5G846082_T01, GRMZM2G068519_T01 и GRMZM2G099305_T01) не было никаких аннотаций на основе результатов BLAST, пять повышался новый изоформ (GRMZM2G025322_T01, GRMZM2G140355_T01, GRMZM2G154580_T01, AC233865.1_FGT001 и GRMZM2G103250_T01) были связаны с реакцией на стресс как биологический процесс, а четыре изоформы с повышенной экспрессией (GRMZM2G140355_T01, GRMZM2G061419_T01, AC233865.1_FGT001 и GRMZM2G106945_T01) были связаны со связыванием ДНК и белков (согласно связыванию с ДНК и белками). Было идентифицировано несколько терминов GO, связанных с транскриптом, которые были связаны с конкретными репродуктивными стадиями, включая опыление, развитие цветка и постэмбриональное развитие в категории биологических процессов (таблица S4).
Термины GO биологического процесса, связанные с дифференциально экспрессируемыми изоформами, включают ответ на стресс, ответ на абиотический стимул и ответ на эндогенный стимул. Мы также идентифицировали несколько новых изоформ транскриптов, индуцированных стрессом. Из 14 новых изоформ, выставляемых значительно другое выражение, изоформы шести транскриптов (GRMZM2G162598_T01, GRMZM2G045239_T01, GRMZM2G163809_T02, GRMZM5G846082_T01, GRMZM2G068519_T01 и GRMZM2G099305_T01) не было никаких аннотаций на основе результатов BLAST, пять повышался новый изоформ (GRMZM2G025322_T01, GRMZM2G140355_T01, GRMZM2G154580_T01, AC233865.1_FGT001 и GRMZM2G103250_T01) были связаны с реакцией на стресс как биологический процесс, а четыре изоформы с повышенной экспрессией (GRMZM2G140355_T01, GRMZM2G061419_T01, AC233865.1_FGT001 и GRMZM2G106945_T01) были связаны со связыванием ДНК и белков (согласно связыванию с ДНК и белками). Было идентифицировано несколько терминов GO, связанных с транскриптом, которые были связаны с конкретными репродуктивными стадиями, включая опыление, развитие цветка и постэмбриональное развитие в категории биологических процессов (таблица S4). Всего было обнаружено, что в 13 транскриптах есть термины GO, относящиеся к репродуктивным стадиям. Двенадцать из этих генов также были идентифицированы как DEG. Исключением был GRMZM2G140355, который проявлял только изменения, специфичные для изоформ.
Всего было обнаружено, что в 13 транскриптах есть термины GO, относящиеся к репродуктивным стадиям. Двенадцать из этих генов также были идентифицированы как DEG. Исключением был GRMZM2G140355, который проявлял только изменения, специфичные для изоформ.
Транскриптомные изменения в чувствительных к засухе генах, связанные со временем цветения
В различных исследованиях сообщалось о генах, играющих роль в регуляции времени цветения. Недавно Li et al. сообщили о 919 генах-кандидатах, связанных со временем цветения кукурузы.на основе анализа опубликованных данных (Li et al., 2016). Гены-кандидаты содержали такие гены, как ZmCCT и ZCN8, которые являются хорошо известными регуляторами времени цветения кукурузы (Meng et al., 2011; Hung et al., 2012) и гомологами многих генов, которые, как было показано, регулируют время цветения кукурузы. другие растения, такие как CONSTANS (CO), FLOWERING LOCUS T (FT), MADS-box и EARLY FLOWERING (Dong et al., 2012; Hung et al., 2012). Чтобы оценить изменения в этих ранее зарегистрированных генах времени цветения в условиях засухи, мы сравнили данные экспрессии этих генов у растений репродуктивной стадии WW и в условиях DS (таблица S4). Всего в обоих условиях экспрессируется 792 гена, а 172 гена не экспрессируются. Из 792 генов только 19 претерпели существенные изменения в условиях засухи. Тринадцать генов (GRMZM2G047055, GRMZM2G154580, GRMZM2G062458, AC208915.3_FG010, GRMZM2G104549, GRMZM2G176173, GRMZM2G137046, GRMZM2G389155, GRMZM2G069146, GRMZM2G005459, GRMZM2G134941, GRMZM2G004483 и GRMZM2G0
Всего в обоих условиях экспрессируется 792 гена, а 172 гена не экспрессируются. Из 792 генов только 19 претерпели существенные изменения в условиях засухи. Тринадцать генов (GRMZM2G047055, GRMZM2G154580, GRMZM2G062458, AC208915.3_FG010, GRMZM2G104549, GRMZM2G176173, GRMZM2G137046, GRMZM2G389155, GRMZM2G069146, GRMZM2G005459, GRMZM2G134941, GRMZM2G004483 и GRMZM2G0
) были повышалась, и шесть генов (GRMZM2G414192, GRMZM2G021777, GRMZM2G012717, GRMZM2G161680, GRMZM2G107886 и GRMZM2G142718) были подавлены (таблица S1).Мы также можем найти гомологи генов из модельных организмов (таблица S5). Мы обнаружили шесть генов со значительными изменениями в экспрессии изоформ (GRMZM2G137046, GRMZM2G004483, GRMZM2G154580, GRMZM2G047055, GRMZM2G140355 и GRMZM2G021777). Ген GRMZM2G140355 не проявлялся статистически дифференциально, но имел значительные дифференциальные транскрипционные изменения. Было обнаружено, что новые транскрипты представляют собой значительно экспрессированные изоформы двух генов (GRMZ2G004483 и GRMZM2G154580) из списка (таблица S2).
qRT-PCR Валидация дифференциально экспрессируемых транскриптов
Чтобы подтвердить точность результатов секвенирования РНК, экспрессию 19 DEG и одно изменение изоформы анализировали с помощью qRT-PCR для трех биологических повторов (рис. 3). За исключением одного гена (GRMZM2G140355), который имеет только одну изоформу со значительно отличающейся экспрессией, все гены были идентифицированы только как DEG. Корреляцию между данными секвенирования РНК и qRT-PCR оценивали путем сравнения кратности изменений в экспрессии генов.Результаты qRT-PCR показали, что картина экспрессии этих генов аналогична наблюдаемой в анализе RNA-seq.
Рисунок 3. Подтверждение относительной экспрессии генов, полученной из секвенирования РНК методом qRT-PCR . Относительные значения экспрессии qRT-PCR представлены как среднее значение SD для трех биологических повторов.
Для подтверждения результатов альтернативного сплайсинга шесть чувствительных к засухе изоформ (GRMZM2G137046, GRMZM2G004483, GRMZM2G154580, GRMZM2G047055, GRMZM2G140355 и GRMZM2G021777) были проверены с использованием количественной ПЦР в реальном времени. Праймеры были разработаны для амплификации определенной области различия между изоформами, и результаты этого анализа показали, что экспрессия изоформы реагировала на засуху (рис. S2–S7), что подтверждает результаты RNA-seq.
Праймеры были разработаны для амплификации определенной области различия между изоформами, и результаты этого анализа показали, что экспрессия изоформы реагировала на засуху (рис. S2–S7), что подтверждает результаты RNA-seq.
Обсуждение
Различные экологические стрессы, такие как засуха, могут прямо или косвенно влиять на производство кукурузы. На протяжении многих лет были предприняты многочисленные попытки выяснить взаимосвязь между временем цветения и стрессом от засухи путем анализа QTL, генетической изменчивости и полиморфизма (Bruce et al., 2002; Фрэнкс и др., 2007 г.; Су и др., 2013; Зийомо и Бернардо, 2013 г.; Сюй и др., 2014; Ли и др., 2016). Что касается времени цветения, ключевыми событиями являются дни до цветения и шелушения, и засуха может увеличить интервал между ними. Повышенный ASI связан с низким уровнем внесения удобрений, в результате чего ASI используется в качестве индикатора устойчивости к стрессу от засухи. Чтобы получить представление о молекулярных событиях, которые регулируют время цветения в ответ на стресс от засухи, мы использовали RNA-seq. По нашим данным, RNA-seq идентифицировал большое количество дифференциально экспрессируемых изоформ транскриптов и новых транскриптов. Поэтому, чтобы глубже понять влияние стресса, вызванного засухой, на ASI, мы исследовали изменения в экспрессии генов и транскриптов в условиях засухи.
По нашим данным, RNA-seq идентифицировал большое количество дифференциально экспрессируемых изоформ транскриптов и новых транскриптов. Поэтому, чтобы глубже понять влияние стресса, вызванного засухой, на ASI, мы исследовали изменения в экспрессии генов и транскриптов в условиях засухи.
Время цветения очень чувствительно к условиям засухи, но широко известно, что сорт кукурузы B73 демонстрирует меньшее изменение ASI в ответ на засуху (Sari-Gorla et al., 1999; Ziyomo and Bernardo, 2013).В среднем АСИ у наших растений в условиях WW имеет продолжительность 1 сут, а в условиях DS увеличивается до 4 сут. Мы находим, что количество дней до отшелушивания не меняется в ответ на засуху, но сдвиг на 3–4 дня происходит из-за опережающего цветения в условиях DS. Растения могут увеличивать время цветения, чтобы повысить выживаемость и обеспечить производство потомства в условиях засушливого стресса (Su et al., 2013; Kooyers, 2015). В этом исследовании мы сосредоточимся на изменениях генов, реагирующих на засуху, среди генов времени цветения в условиях короткого дня. Как известно, для В73 характерны растения умеренного климата, адаптированные к условиям длинного дня. В нашем исследовании есть два важных фактора окружающей среды, такие как засуха и фотопериод, которые могут влиять на гены времени цветения. Учитывая условия окружающей среды, мы провели анализ с высокой отсечкой для большей точности и получили ДЭГ и изоформы.
Как известно, для В73 характерны растения умеренного климата, адаптированные к условиям длинного дня. В нашем исследовании есть два важных фактора окружающей среды, такие как засуха и фотопериод, которые могут влиять на гены времени цветения. Учитывая условия окружающей среды, мы провели анализ с высокой отсечкой для большей точности и получили ДЭГ и изоформы.
Было показано, что время цветения напрямую связано с урожайностью зерна, поэтому во многих исследованиях изучалось время цветения злаковых и модельных растений (Buckler et al., 2009; Юнг и Мюллер, 2009 г.; Конг и др., 2010 г.; Эндо-Хигаси и Идзава, 2011 г.; Чен и др., 2012; Хунг и др., 2012; Ли и др., 2016). На основании этих исследований мы смогли проанализировать экспрессию гомологов генов, о которых ранее сообщалось, что они связаны со временем цветения у других растений (Li et al., 2016). Ранее опубликованные данные экспрессии согласовывались со значениями экспрессии из наших данных РНК-сек (таблица S4). Наши результаты идентифицировали 19 генов со значительно измененными генами, заявленными как кандидаты в регуляторы времени цветения кукурузы. Было подтверждено, что среди этих 19 генов шесть генов (GRMZM2G004483, GRMZM2G154580, GRMZM2G176173, GRMZM2G0
Было подтверждено, что среди этих 19 генов шесть генов (GRMZM2G004483, GRMZM2G154580, GRMZM2G176173, GRMZM2G0
, GRMZM2G140355 и GRMZM2G021777) играют роль в биологических процессах, связанных со временем цветения, таких как размножение, развитие цветка и постэмбриональное развитие (рис. 2). . Таким образом, изменения в экспрессии этих генов могут непосредственно приводить к изменению продолжительности цветения в условиях засухи.
Было обнаружено, что подмножество идентифицированных нами генов, реагирующих на засуху, обладает доменами CCT [CONSTANS, CONSTANS-LIKE и ВРЕМЯ СВЯЗЫВАНИЯ ХЛОРОФИЛЛА A/B1 (TOC1)].Было показано, что белки, содержащие домен CCT, играют роль в регуляции времени цветения (Xue et al., 2008; Hung et al., 2012) и подвержены стрессу от засухи (Weng et al., 2014). Было обнаружено, что два гена (GRMZM2G09236 и GRMZM2G176173), сходные с CONSTANS CO8, активируются в условиях засухи. Сообщалось, что гены CONSTANS CO8 играют ту же роль, что и гены числа зерен, высоты растений и даты колошения7 (Ghd7), которые считаются регуляторами даты колошения и потенциальной урожайности сельскохозяйственных культур (Coelho et al. , 2013; Венг и др., 2014). Напротив, гены (GRMZM2G021777 и GRMZM2G107886), связанные с CONSTANS CO5, и ген, содержащий цинковые пальцы, CONSTANS-LIKE 16, были подавлены. CONSTANS CO5 имеет сходство с белком цинковых пальцев CONSTANS-LIKE 3 (COL3), который был идентифицирован как взаимодействующий с COP1 у Arabidopsis (Datta, 2006). Было показано, что COP1 регулирует время цветения, действуя как репрессор цветения (Reed et al., 1994). Гены, связанные с TOC1, показали противоположные ответы. Два гена (AC208915.3_FG010 и GRMZM2G104549) активировались, но третий (GRMZM2G414192) подавлялся в условиях засухи. Сообщалось, что эти белки, содержащие домен CCT, участвуют в цветении и регуляции циркадных ритмов посредством активации SUPPRESSOR OF CONSTANS (SOC1) (Samach, 2000; Wenkel et al., 2006). Над доменами CCT находится ген, принадлежащий к семейству Dof (GRMZM2G142718), который связан с контролем времени цветения (Kim et al., 2006; Fornara et al., 2009; Corrales et al., 2014) также был идентифицирован как ген с пониженной экспрессией.
, 2013; Венг и др., 2014). Напротив, гены (GRMZM2G021777 и GRMZM2G107886), связанные с CONSTANS CO5, и ген, содержащий цинковые пальцы, CONSTANS-LIKE 16, были подавлены. CONSTANS CO5 имеет сходство с белком цинковых пальцев CONSTANS-LIKE 3 (COL3), который был идентифицирован как взаимодействующий с COP1 у Arabidopsis (Datta, 2006). Было показано, что COP1 регулирует время цветения, действуя как репрессор цветения (Reed et al., 1994). Гены, связанные с TOC1, показали противоположные ответы. Два гена (AC208915.3_FG010 и GRMZM2G104549) активировались, но третий (GRMZM2G414192) подавлялся в условиях засухи. Сообщалось, что эти белки, содержащие домен CCT, участвуют в цветении и регуляции циркадных ритмов посредством активации SUPPRESSOR OF CONSTANS (SOC1) (Samach, 2000; Wenkel et al., 2006). Над доменами CCT находится ген, принадлежащий к семейству Dof (GRMZM2G142718), который связан с контролем времени цветения (Kim et al., 2006; Fornara et al., 2009; Corrales et al., 2014) также был идентифицирован как ген с пониженной экспрессией.
Мы также наблюдали дифференциальную экспрессию изоформ для подмножества генов-кандидатов (GRMZM2G137046, GRMZM2G004483, GRMZM2G154580, GRMZM2G047055, GRMZM2G140355 и GRMZM2G021777). Ген GRMZM2G140355 не экспрессировался статистически дифференциально, но проявлял изменения в экспрессии изоформы. Новые транскрипты были идентифицированы для двух из этих генов (GRMZM2G140355 и GRMZM2G154580). Предполагается, что новый транскрипт для GRMZM2G140355 связан с фактором транскрипции HY5 в соответствии с результатами BLAST, который, как описано выше, взаимодействует непосредственно с COP1 (Reyes et al., 2004). Другой новый транскрипт (GRMZM2G154580) имеет бластные результаты, сходные с PRR37, который, как сообщалось, действует как цветочный репрессор FT-подобных генов, которые задерживают время цветения у сорго (Johnson et al., 1994b).
В ответ на засуху альтернативный сплайсинг генов, описанных выше, усиливал экспрессию функциональной изоформы. Альтернативный сплайсинг увеличивает функциональную способность генов и дает возможность для регуляции генов и выполнения других функций (Yan et al. , 2012). Как сделать более одного транскрипта мРНК из транскриптов пре-мРНК. Возможны дифференциальные сайты сплайсинга, которые определяются взаимодействием связывающих белков, факторов транскрипта и факторов сплайсинга, которые направляют сплайсосому (Nilsen and Graveley, 2010; Wachter et al., 2012). Консенсусные последовательности изоформ, полученные в наших результатах (таблица S2), также могут улучшить их функции, скорость реакции и дать новые функции.
, 2012). Как сделать более одного транскрипта мРНК из транскриптов пре-мРНК. Возможны дифференциальные сайты сплайсинга, которые определяются взаимодействием связывающих белков, факторов транскрипта и факторов сплайсинга, которые направляют сплайсосому (Nilsen and Graveley, 2010; Wachter et al., 2012). Консенсусные последовательности изоформ, полученные в наших результатах (таблица S2), также могут улучшить их функции, скорость реакции и дать новые функции.
В дополнение к этим генам-кандидатам мы также идентифицировали гены, которые, по-видимому, связаны с изменениями времени цветения на основе результатов BLAST.Несколько генов, отвечающих за время цветения, содержали домены ССТ и были обнаружены в списке DEG. Среди генов с пониженной экспрессией GRMZM2G012717 родственен гену, содержащему цинковые пальцы, CONSTANS-LIKE 16 (описанному выше). Фактор транскрипции GATA (GRMZM2G039586), играющий сходную с CONSTANS роль, был обнаружен в активированных генах (Reyes, 2004). Однако FLOWERING LOCUS C (FLC), репрессор транскрипции SOC1, не был идентифицирован как DEG. В дополнение к генам, содержащим домен CCT, было обнаружено, что другие гены, связанные со временем цветения, также по-разному регулируются.Субстрат фитохромкиназы 1 (PKS1; GRMZM2G066291), который играет роль в стимуляции цветения (Johnson et al., 1994b; Reed et al., 1994), активировался. C 2 H 2 Ген цинкового пальца (GRMZM2G105224), родственный генам, контролирующим время цветения (Kim et al., 2006; Fornara et al., 2009; Corrales et al., 2014), обнаружен у подавленные гены (таблица S1). Ген (GRMZM2G171912), родственный HY5 (описан выше), и два гена (GRMZM2G079632 и GRMZM2G159500) с доменами NAC проявляли дифференциальную изоформную экспрессию.Известно, что белки NAC-домена контролируют реакции развития на стресс, включая время цветения (Olsen et al., 2005; Yoo et al., 2007). Было показано, что транскрипционные факторы bHLH контролируют множество соответствующих процессов, включая время цветения и абиотический стресс (Ito et al., 2012; Liu et al., 2013). В нашем списке два гена (GRMZM2G350312 и GRMZM2G005939) были аннотированы как bHLH-подобные транскрипты.
В дополнение к генам, содержащим домен CCT, было обнаружено, что другие гены, связанные со временем цветения, также по-разному регулируются.Субстрат фитохромкиназы 1 (PKS1; GRMZM2G066291), который играет роль в стимуляции цветения (Johnson et al., 1994b; Reed et al., 1994), активировался. C 2 H 2 Ген цинкового пальца (GRMZM2G105224), родственный генам, контролирующим время цветения (Kim et al., 2006; Fornara et al., 2009; Corrales et al., 2014), обнаружен у подавленные гены (таблица S1). Ген (GRMZM2G171912), родственный HY5 (описан выше), и два гена (GRMZM2G079632 и GRMZM2G159500) с доменами NAC проявляли дифференциальную изоформную экспрессию.Известно, что белки NAC-домена контролируют реакции развития на стресс, включая время цветения (Olsen et al., 2005; Yoo et al., 2007). Было показано, что транскрипционные факторы bHLH контролируют множество соответствующих процессов, включая время цветения и абиотический стресс (Ito et al., 2012; Liu et al., 2013). В нашем списке два гена (GRMZM2G350312 и GRMZM2G005939) были аннотированы как bHLH-подобные транскрипты. Два подавленных гена (GRMZM2G105224 и GRMZM5G801627) содержали C 2 H 2 и цинковые пальцы CCCH.Сообщалось, что аналогичные белки цинковых пальцев играют роль в подавлении времени цветения (Weingartner et al., 2011; Chao et al., 2014). Кроме того, уже сообщалось, что белки, содержащие домен C 2 H 2 , bHLH и NAC, важны для ответа на различные абиотические стрессы (Shinozaki and Yamaguchi-Shinozaki, 2007; Marino et al., 2008; Golldack). и др., 2014). Было высказано предположение, что некоторые из этих генов, участвующих в путях реакции на абиотический стресс, могут изменять время цветения растений.
Два подавленных гена (GRMZM2G105224 и GRMZM5G801627) содержали C 2 H 2 и цинковые пальцы CCCH.Сообщалось, что аналогичные белки цинковых пальцев играют роль в подавлении времени цветения (Weingartner et al., 2011; Chao et al., 2014). Кроме того, уже сообщалось, что белки, содержащие домен C 2 H 2 , bHLH и NAC, важны для ответа на различные абиотические стрессы (Shinozaki and Yamaguchi-Shinozaki, 2007; Marino et al., 2008; Golldack). и др., 2014). Было высказано предположение, что некоторые из этих генов, участвующих в путях реакции на абиотический стресс, могут изменять время цветения растений.
Сообщалось об альтернативных событиях сплайсинга, вызванных абиотическим стрессом, у растений (Li et al., 2013; Staiger and Brown, 2013; Thatcher et al., 2016). Эти типы изменений позволяют растениям быстрее реагировать на изменения окружающей среды и могут играть важную роль в выживании и производстве потомства (Sonenberg and Hinnebusch, 2009; Lackner et al. , 2012). Наши результаты выявили 617 чувствительных к засухе DEG и 126 изоформных изменений, связанных со временем цветения в условиях засушливого стресса. Среди генов-кандидатов, о которых ранее сообщалось, что они связаны со временем цветения, мы подтвердили, что 19 являются генами, реагирующими на засуху.В дополнение к этим генам-кандидатам мы также определили новые гены, которые потенциально могут быть вовлечены во время цветения на основе результатов BLAST. Было обнаружено, что некоторые факторы транскрипции (HY5 и PRR37), которые действуют как репрессоры белков, которые могут играть роль в задержке времени цветения, активируются. Точно так же известные репрессоры времени цветения (содержащие C 2 H 2 и домены цинковых пальцев CCCH) подавлялись.
, 2012). Наши результаты выявили 617 чувствительных к засухе DEG и 126 изоформных изменений, связанных со временем цветения в условиях засушливого стресса. Среди генов-кандидатов, о которых ранее сообщалось, что они связаны со временем цветения, мы подтвердили, что 19 являются генами, реагирующими на засуху.В дополнение к этим генам-кандидатам мы также определили новые гены, которые потенциально могут быть вовлечены во время цветения на основе результатов BLAST. Было обнаружено, что некоторые факторы транскрипции (HY5 и PRR37), которые действуют как репрессоры белков, которые могут играть роль в задержке времени цветения, активируются. Точно так же известные репрессоры времени цветения (содержащие C 2 H 2 и домены цинковых пальцев CCCH) подавлялись.
Таким образом, мы предполагаем, что изменения в экспрессии этих генов, реагирующих на засуху, способствуют цветению, что приводит к общему увеличению ASI.В целом, наши результаты обеспечивают полногеномный анализ DEG, новых транскриптов и изменений экспрессии изоформ на репродуктивной стадии кукурузы в условиях засушливого стресса. Дальнейшая характеристика этих изменений генетической регуляции будет иметь большое значение для улучшения селекции кукурузы.
Дальнейшая характеристика этих изменений генетической регуляции будет иметь большое значение для улучшения селекции кукурузы.
Выводы
В настоящем исследовании мы исследовали изменения транскрипции в ответ на стресс от засухи во время цветения кукурузы в условиях короткого дня. Используя RNA-seq, мы проанализировали изменения транскрипции, специфичной для генов и изоформ, чтобы идентифицировать гены, участвующие во времени цветения в условиях засушливого стресса.Эти гены, вероятно, прямо или косвенно связаны с изменениями в ASI у кукурузы. Дальнейшая характеристика этих изменений транскриптома улучшит наше понимание регуляции процессов, связанных со временем цветения в условиях засухи.
Вклад авторов
KS и HK разработали и провели эксперимент, а KS провел работу по биоинформатике и написал рукопись. HK, SS и KK провели полевые эксперименты и отбор проб. JM и JK помогают в работе по биоинформатике.BL консультировал по экспериментам и анализу данных. Все авторы обсудили результаты и одобрили рукопись.
Все авторы обсудили результаты и одобрили рукопись.
Заявление о конфликте интересов
Авторы заявляют, что исследование проводилось при отсутствии каких-либо коммерческих или финансовых отношений, которые могли бы быть истолкованы как потенциальный конфликт интересов.
Благодарности
Работа выполнена при поддержке «Совместной исследовательской программы развития сельскохозяйственной науки и техники» (Проект №PJ0126417)» Управление по развитию сельских районов, Республика Корея. Это исследование было поддержано 213009-05-1-SB710, Министерством сельского хозяйства, продовольствия и сельских районов (MAFRA), Министерством океанов и рыболовства (MOF), Управлением сельского развития (RDA) и Корейской лесной службой (KFS).
Дополнительный материал
Дополнительный материал к этой статье можно найти в Интернете по адресу: https://www.frontiersin.org/article/10.3389/fpls.2017.00267/full#supplementary-material
.Наличие вспомогательных данных
Наборы данных, поддерживающие результаты этой статьи, доступны в репозитории NCBI Sequence Read Archive (SRA), [Http://www. ncbi.nlm.nih.gov/sra?term=SRP082534].
ncbi.nlm.nih.gov/sra?term=SRP082534].
Рисунок S1. Относительная экспрессия ZmDREB2A в качестве маркеров стресса от засухи . Относительные значения экспрессии qRT-PCR представлены как среднее стандартное отклонение трех биологических повторностей.
Рисунок S2. qRT-PCR подтверждение относительной экспрессии изоформы из чувствительного к засухе гена (GRMZM2G004483). (A) График Сашими гена под WW, (B) График Сашими гена под DS, (C) синие дорожки показывают охват чтения РНК-секвенции, а аннотированные гены и красные дорожки показывают подтвержденную область с использованием qRT-PCR , (D) данные относительной экспрессии (log 2 кратное изменение) из qRT-PCR при каждом условии.Относительные значения экспрессии qRT-PCR представлены как среднее стандартное отклонение трех биологических повторностей.
Рисунок S3. qRT-PCR подтверждение относительной экспрессии изоформы из чувствительного к засухе гена (GRMZM2G021777). (A) График Сашими гена под WW, (B) График Сашими гена под DS, (C) синие дорожки показывают охват чтения РНК-секвенции, а аннотированные гены и красные дорожки показывают подтвержденную область с использованием qRT-PCR , (D) данные относительной экспрессии (log 2 кратное изменение) из qRT-PCR при каждом условии.Относительные значения экспрессии qRT-PCR представлены как среднее стандартное отклонение трех биологических повторностей.
(A) График Сашими гена под WW, (B) График Сашими гена под DS, (C) синие дорожки показывают охват чтения РНК-секвенции, а аннотированные гены и красные дорожки показывают подтвержденную область с использованием qRT-PCR , (D) данные относительной экспрессии (log 2 кратное изменение) из qRT-PCR при каждом условии.Относительные значения экспрессии qRT-PCR представлены как среднее стандартное отклонение трех биологических повторностей.
Рисунок S4. qRT-PCR подтверждение относительной экспрессии изоформы из чувствительного к засухе гена (GRMZM2G047055). (A) График Сашими гена под WW, (B) График Сашими гена под DS, (C) синие дорожки показывают охват чтения РНК-секвенции, а аннотированные гены и красные дорожки показывают подтвержденную область с использованием qRT-PCR , (D) данные относительной экспрессии (log 2 кратное изменение) из qRT-PCR при каждом условии. Относительные значения экспрессии qRT-PCR представлены как среднее стандартное отклонение трех биологических повторностей.
Относительные значения экспрессии qRT-PCR представлены как среднее стандартное отклонение трех биологических повторностей.
Рисунок S5. qRT-PCR подтверждение относительной экспрессии изоформы из чувствительного к засухе гена (GRMZM2G137046). (A) График Сашими гена под WW, (B) График Сашими гена под DS, (C) синие дорожки показывают охват чтения РНК-секвенции, а аннотированные гены и красные дорожки показывают подтвержденную область с использованием qRT-PCR , (D) данные относительной экспрессии (log 2 кратное изменение) из qRT-PCR при каждом условии.Относительные значения экспрессии qRT-PCR представлены как среднее стандартное отклонение трех биологических повторностей.
Рисунок S6. qRT-PCR подтверждение относительной экспрессии изоформы из чувствительного к засухе гена (GRMZM2G140355). (A) График Сашими гена под WW, (B) График Сашими гена под DS, (C) синие дорожки показывают охват чтения РНК-секвенции, а аннотированные гены и красные дорожки показывают подтвержденную область с использованием qRT-PCR , (D) данные относительной экспрессии (log 2 кратное изменение) из qRT-PCR при каждом условии.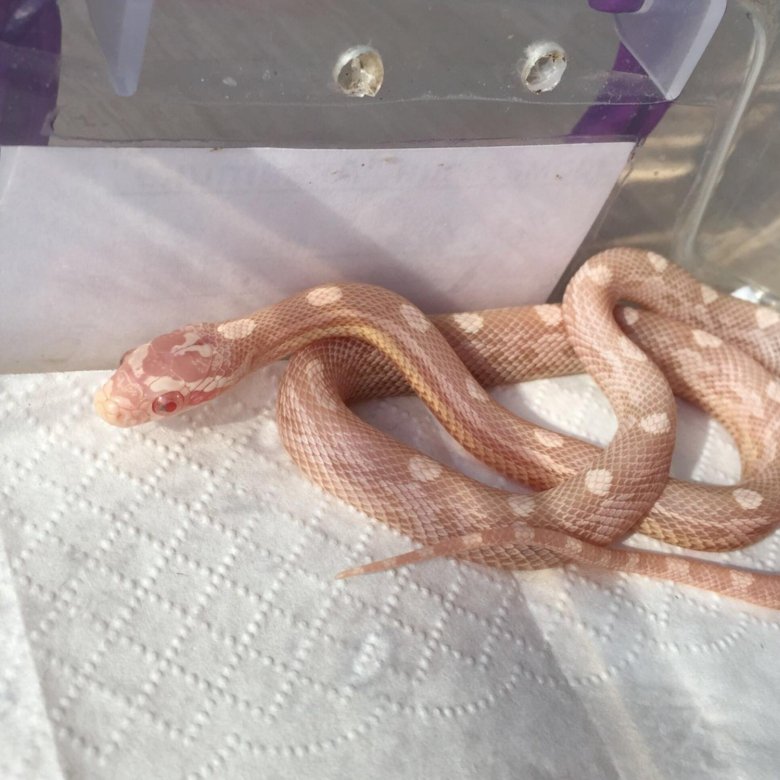 Относительные значения экспрессии qRT-PCR представлены как среднее стандартное отклонение трех биологических повторностей.
Относительные значения экспрессии qRT-PCR представлены как среднее стандартное отклонение трех биологических повторностей.
Рисунок S7. qRT-PCR подтверждение относительной экспрессии изоформы из чувствительного к засухе гена (GRMZM2G154580). (A) График Сашими гена под WW, (B) График Сашими гена под DS, (C) синие дорожки показывают охват чтения РНК-секвенции, а аннотированные гены и красные дорожки показывают подтвержденную область с использованием qRT-PCR , (D) данные относительной экспрессии (log 2 кратное изменение) из qRT-PCR при каждом условии.Относительные значения экспрессии qRT-PCR представлены как среднее стандартное отклонение трех биологических повторностей.
Таблица S1. Список дифференциально экспрессируемых генов (DEG) при засухе .
Таблица S2. Список дифференциально экспрессируемых изоформ .
Таблица S3. Условия GO Slim по результатам BiNGO .
Таблица S4. Значения экспрессии генов-кандидатов времени цветения из опубликованных данных .
Таблица S5. Список генов, реагирующих на засуху, участвующих во времени цветения .
Ссылки
Андраде Ф.Х., Вега К., Ухарт С., Сирило А., Кантареро М. и Валентинуз О. (1999). Определение числа зерен кукурузы. Растениеводство. 39, 453. doi: 10.2135/cropsci1999.0011183X003
00026x
Полнотекстовая перекрестная ссылка | Академия Google
Банерджи, А., и Ройчаудхури, А. (2015). Белки WRKY: передача сигналов и регуляция экспрессии во время реакции на абиотический стресс. науч. Мир J. 2015, 1–17. дои: 10.1155/2015/807560
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Бенджамини, Ю.и Екутиэли, Д. (2001). Контроль частоты ложных обнаружений при многократном тестировании в зависимости. Энн. Стат. 1, 1165–1188. дои: 10.2307/2674075
Полнотекстовая перекрестная ссылка | Академия Google
Берналь, М. , Эстиарте, М., и Пеньюэлас, Дж. (2011). Засуха опережает фенологию весеннего роста средиземноморского кустарника Erica multiflora . Растение Биол. 13, 252–257. doi: 10.1111/j.1438-8677.2010.00358.x
, Эстиарте, М., и Пеньюэлас, Дж. (2011). Засуха опережает фенологию весеннего роста средиземноморского кустарника Erica multiflora . Растение Биол. 13, 252–257. doi: 10.1111/j.1438-8677.2010.00358.x
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Блюм, А.(1996). Реакция культур на засуху и интерпретация адаптации. Регулятор роста растений. 20, 135–148. дои: 10.1007/BF00024010
Полнотекстовая перекрестная ссылка | Академия Google
Боланьос, Дж., и Эдмидес, Г. О. (1996). Важность интервала цветения-шелушения в селекции на засухоустойчивость тропической кукурузы. Полевые культуры Res. 48, 65–80.
Академия Google
Брюс, В. Б., Эдмидес, Г. О., и Баркер, Т. С. (2002). Молекулярные и физиологические подходы к улучшению засухоустойчивости кукурузы. Дж. Экспл. Бот. 53, 13–25. doi: 10.1093/jxb/53.366.13
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Buckler, E. S., Holland, J.B., Bradbury, P.J., Acharya, C.B., Brown, P.J., Browne, C., et al. (2009). Генетическая архитектура времени цветения кукурузы. Наука 325, 714–718. doi: 10.1126/science.1174276
S., Holland, J.B., Bradbury, P.J., Acharya, C.B., Brown, P.J., Browne, C., et al. (2009). Генетическая архитектура времени цветения кукурузы. Наука 325, 714–718. doi: 10.1126/science.1174276
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Бирн, П. Ф., Боланьос, Дж., Эдмидес, Г. О., и Итон, Д.Л. (1995). Преимущества селекции в условиях засухи по сравнению с тестированием в нескольких местах в родственных тропических популяциях кукурузы. Растениеводство. 35, 63.
Академия Google
Каттивелли Л., Рицца Ф., Бадек Ф.-В., Маццукотелли Э., Мастранжело А.М., Франсия Э. и др. (2008). Улучшение засухоустойчивости сельскохозяйственных культур: комплексный подход от селекции к геномике. Полевые культуры Res. 105, 1–14. doi: 10.1016/j.fcr.2007.07.004
Полнотекстовая перекрестная ссылка | Академия Google
Чао Ю., Zhang, T., Yang, Q., Kang, J., Sun, Y., Gruber, M.Y., et al. (2014). Экспрессия гена белка цинковых пальцев CCCH-типа люцерны MsZFN задерживает время цветения у трансгенных Arabidopsis thaliana .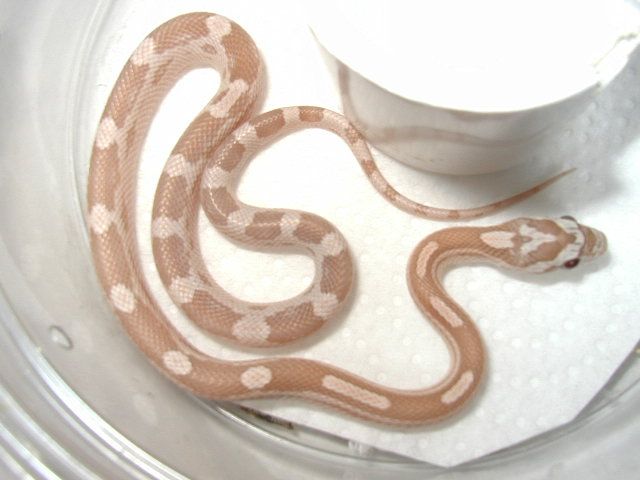 Растениевод. 215–216, 92–99. doi: 10.1016/j.plantsci.2013.10.012
Растениевод. 215–216, 92–99. doi: 10.1016/j.plantsci.2013.10.012
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Чен К., ДеКлерк Г., Тиан Ф., Спунер В., Маккауч С. и Баклер Э. (2012). PICARA, аналитический конвейер, обеспечивающий вероятностный вывод о априорных генов-кандидатов, лежащих в основе общегеномных ассоциаций QTL у растений. PLoS ONE 7:e46596. doi: 10.1371/journal.pone.0046596
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Коэльо, К.П., Коста Нетто, А.П., Коласанти, Дж., и Чалфун-Джуниор, А. (2013). Предлагаемая модель сигнального пути цветения сахарного тростника под фотопериодическим контролем. Жен. Мол. Рез. 12, 1347–1359. doi: 10.4238/2013.April.25.6
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Корралес, А. Р., Nebauer, S.G., Carrillo, L., Fernandez-Nohales, P., Marques, J., Renau-Morata, B., et al. (2014). Характеристика циклических факторов Dof томата выявляет сохранившиеся и новые функции в контроле времени цветения и реакции на абиотический стресс. Дж. Экспл. Бот. 65, 995–1012. doi: 10.1093/jxb/ert451
Дж. Экспл. Бот. 65, 995–1012. doi: 10.1093/jxb/ert451
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Донг З., Данилевская О., Абади Т., Мессина К., Коулз Н. и Купер М. (2012). Модель генной регуляторной сети цветочного перехода верхушки побега кукурузы и ее динамическое моделирование. PLoS ONE 7:e43450. doi: 10.1371/journal.pone.0043450
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Edmeades, G.O., Bänziger, M., Elings, A., Chapman, S.C., and Ribaut, J.M. (1997). «Последние достижения в селекции кукурузы на засухоустойчивость», в Applications of Systems Approaches on the Field Level , eds MJ Kropff, PS Teng, PK Aggarwal, J. Bouma, BAM Bouman, JW Jones и HH van Laar (Dordrecht : Спрингер), 63–78.дои: 10.1007/978-94-017-0754-1_5
Полнотекстовая перекрестная ссылка | Академия Google
Эндо-Хигаси, Н., и Идзава, Т. (2011). Гены времени цветения дата колошения 1 и ранняя дата колошения 1 вместе контролируют развитие метелки у риса. Физиол клеток растений. 52, 1083–1094. doi: 10.1093/pcp/pcr059
Физиол клеток растений. 52, 1083–1094. doi: 10.1093/pcp/pcr059
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Филичкин С.А., Прист Х.Д., Гиван С.А., Шен Р., Брайант Д.В., Фокс С.Е. и др. (2010).Полногеномное картирование альтернативного сплайсинга у Arabidopsis thaliana . Рез. генома. 20, 45–58. doi: 10.1101/gr.0
.109Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Fornara, F., Panigrahi, K.C., Gissot, L., Sauerbrunn, N., Rühl, M., Jarillo, J.A., et al. (2009). Факторы транскрипции DOF арабидопсиса действуют избыточно, снижая экспрессию CONSTANS, и необходимы для реакции фотопериодического цветения. Дев. Ячейка 17, 75–86.doi: 10.1016/j.devcel.2009.06.015
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Фрэнкс, С.Дж., Сим, С., и Вейс, А.Е. (2007). Быстрая эволюция времени цветения однолетних растений в ответ на колебания климата. Проц. Натл. акад. науч. США 104, 1278–1282. doi: 10.1073/pnas.0608379104
Проц. Натл. акад. науч. США 104, 1278–1282. doi: 10.1073/pnas.0608379104
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Фрей, Ф. П., Урбани, К., Хюттель, Б., Райнхардт, Р., и Стич, Б. (2015).Полногеномное профилирование экспрессии и фенотипическая оценка инбредных сортов европейской кукурузы на стадии проростков в ответ на тепловой стресс. BMC Genomics 16:123. doi: 10.1186/s12864-015-1282-1
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Гарг Р., Шанкар Р., Таккар Б., Кудапа Х., Кришнамурти Л., Мантри Н. и др. (2016). Анализ транскриптома выявляет специфические для генотипа и стадии развития молекулярные реакции на засуху и солевой стресс у нута. науч. Респ. 6:19228. дои: 10.1038/srep19228
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Голлдак, Д., Ли, К., Мохан, Х., и Пробст, Н. (2014). Устойчивость растений к засухе и солевому стрессу: распутывание сигнальных сетей. Фронт. Растениевод. 5:151. doi: 10.3389/fpls.2014.00151
Фронт. Растениевод. 5:151. doi: 10.3389/fpls.2014.00151
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Холл, А. Дж., Чименти, К., Трапани, Н., Вилелла, Ф., и де Унау, Р.С. (1984). Урожайность генотипов кукурузы, подверженных водному стрессу: связь с признаками, измеренными у проростков и цветущих растений. Полевые культуры Res. 9, 41–57. дои: 10.1016/0378-4290(84)-4
Полнотекстовая перекрестная ссылка | Академия Google
Hansey, C.N., Vaillancourt, B., Sekhon, R.S., de Leon, N., Kaeppler, S.M., and Buell, C.R. (2012). Разнообразие генома кукурузы ( Zea mays L.), выявленное секвенированием РНК. PLoS ONE 7:e33071. doi: 10.1371/журнал.пон.0033071
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Hu, X., Liu, R., Li, Y., Wang, W., Tai, F., Xue, R., et al. (2009). Белок теплового шока 70 регулирует индуцированную абсцизовой кислотой антиоксидантную реакцию кукурузы на комбинированную засуху и тепловой стресс. Регулятор роста растений. 60, 225–235. doi: 10.1007/s10725-009-9436-2
Регулятор роста растений. 60, 225–235. doi: 10.1007/s10725-009-9436-2
Полнотекстовая перекрестная ссылка | Академия Google
Гумберт, С., Субеди, С., Кон, Дж., Зенг, Б., Би, Ю.М., Чен, X., и др.(2013). Профилирование полногеномной экспрессии кукурузы в ответ на индивидуальный и комбинированный стресс воды и азота. BMC Genomics 14:3. дои: 10.1186/1471-2164-14-3
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Hung, H.Y., Shannon, L.M., Tian, F., Bradbury, P.J., Chen, C., Flint-Garcia, S.A., et al. (2012). ZmCCT и генетическая основа адаптации к длине дня, лежащая в основе распространения кукурузы после одомашнивания. Проц. Натл. акад. науч. У.SA 109, E1913–E1921. doi: 10.1073/pnas.1203189109
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Ito, S., Song, Y.H., Josephson-Day, A.R., Miller, R.J., Breton, G., Olmstead, R.G., et al. (2012). Активаторы транскрипции FLOWERING BHLH контролируют экспрессию фотопериодического регулятора цветения CONSTANS у арабидопсиса. Проц. Натл. акад. науч. США 109, 3582–3587. doi: 10.1073/pnas.1118876109
Проц. Натл. акад. науч. США 109, 3582–3587. doi: 10.1073/pnas.1118876109
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Цзян, Т., Fountain, J., Davis, G., Kemerait, R., Scully, B., Lee, R.D., et al. (2012). Морфология корней и анализ экспрессии генов в ответ на стресс от засухи у кукурузы ( Zea mays ). Завод Мол. биол. Респ. 30, 360–369. doi: 10.1007/s11105-011-0347-9
Полнотекстовая перекрестная ссылка | Академия Google
Джонсон, Э., Брэдли, М., Харберд, Н.П., и Уайтлам, Г.К. (1994b). Фотоответы выращенных на свету мутантов phyA арабидопсиса (фитохром А необходим для восприятия удлинения светового дня). Физиол растений. 105, 141–149. doi: 10.1104/стр.105.1.141
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Какуману, А., Амбаварам, М.М., Клумас, К., Кришнан, А., Батланг, У., Майерс, Э., и др. (2012). Влияние засухи на экспрессию генов в репродуктивной и листовой меристемной ткани кукурузы, выявленное с помощью RNA-seq. Физиол растений. 160, 846–867. doi: 10.1104/pp.112.200444
Физиол растений. 160, 846–867. doi: 10.1104/pp.112.200444
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Ким, Д., Pertea, G., Trapnell, C., Pimentel, H., Kelley, R., and Salzberg, S.L. (2013). TopHat2: точное выравнивание транскриптомов при наличии вставок, делеций и слияний генов. Геном Биол. 14:1. doi: 10.1186/gb-2013-14-4-r36
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Ким, С., Чой, К., Пак, К., Хван, Х.Дж., и Ли, И. (2006). СУПРЕСОР FRIGIDA4, кодирующий белок цинковых пальцев типа C2h3, подавляет цветение за счет активации транскрипции Arabidopsis FLOWERING LOCUS C. Растительная клетка 18, 2985–2998. doi: 10.1105/tpc.106.045179
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Kong, F., Liu, B., Xia, Z., Sato, S., Kim, B.M., Watanabe, S., et al. (2010). Два согласованно регулируемых гомолога FLOWERING LOCUS T участвуют в контроле фотопериодического цветения сои. Физиол растений. 154, 1220–1231. doi: 10.1104/стр.110.160796
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Красенский Ю.и Йонак, К. (2012). Метаболические перестройки и регуляторные сети, вызванные засухой, солью и температурным стрессом. Дж. Экспл. Бот. 63, 1593–1608. дои: 10.1093/jxb/err460
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Лакнер, Д. Х., Шмидт, М. В., Ву, С., Вольф, Д. А., и Бахлер, Дж. (2012). Регуляция транскриптома, трансляции и протеома в ответ на стресс окружающей среды у делящихся дрожжей. Геном Биол. 13:R25. doi: 10.1186/gb-2012-13-4-r25
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Ли, В., Лин, В.-Д., Рэй, П., Лан, П., и Шмидт, В. (2013). Полногеномное обнаружение чувствительного к состоянию альтернативного сплайсинга в корнях арабидопсиса. Физиол растений. 162, 1750–1763. doi: 10.1104/стр.113.217778
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Li, Y. X., Li, C., Bradbury, P.J., Liu, X., Lu, F., Romay, C.M., et al. (2016). Идентификация генетических вариантов, связанных со временем цветения кукурузы, с использованием чрезвычайно большой мультигенетической фоновой популяции. Завод J. 86, 391–402. doi: 10.1111/tpj.13174
X., Li, C., Bradbury, P.J., Liu, X., Lu, F., Romay, C.M., et al. (2016). Идентификация генетических вариантов, связанных со временем цветения кукурузы, с использованием чрезвычайно большой мультигенетической фоновой популяции. Завод J. 86, 391–402. doi: 10.1111/tpj.13174
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Лю, Ю., Ли, X., Ли, К., Лю, Х., и Лин, К. (2013). Множественные белки bHLH образуют гетеродимеры, чтобы опосредовать CRY2-зависимую регуляцию времени цветения у арабидопсиса. Генетика PLoS. 9:e1003861. doi: 10.1371/journal.pgen.1003861
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Лю Ю., Чжоу М., Гао З., Рен В., Ян Ф., Хе Х. и др. (2015). Анализ РНК-секвенции выявляет членов семейства MAPKKK, связанных с засухоустойчивостью кукурузы. PLoS ONE 10:e0143128. doi: 10.1371/journal.pone.0143128
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Ливак, К. Дж., и Шмитген, Т. Д. (2001). Анализ данных об относительной экспрессии генов с использованием количественной ПЦР в реальном времени и метода 2 -ΔΔCT . Методы 25, 402–408. doi: 10.1006/meth.2001.1262
Дж., и Шмитген, Т. Д. (2001). Анализ данных об относительной экспрессии генов с использованием количественной ПЦР в реальном времени и метода 2 -ΔΔCT . Методы 25, 402–408. doi: 10.1006/meth.2001.1262
Полнотекстовая перекрестная ссылка | Академия Google
Маэре, С., Хейманс, К., и Куйпер, М. (2005). BiNGO: плагин Cytoscape для оценки чрезмерного представления категорий генной онтологии в биологических сетях. Биоинформатика 21, 3448–3449. doi: 10.1093/биоинформатика/bti551
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Марино, Р., Поннайа, М., Краевски, П., Фрова, К., Джанфранчески, Л., Пе, М.Е., и др. (2008). Решение проблемы засухоустойчивости кукурузы с помощью транскрипционного профилирования и картирования. мол. Жене.Геномика 281, 163–179. doi: 10.1007/s00438-008-0401-y
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
McMullen, M.D., Kresovich, S. , Villeda, H.S., Bradbury, P., Li, H., Sun, Q., et al. (2009). Генетические свойства картографической популяции вложенных ассоциаций кукурузы. Наука 325, 737–740. doi: 10.1126/science.1174320
, Villeda, H.S., Bradbury, P., Li, H., Sun, Q., et al. (2009). Генетические свойства картографической популяции вложенных ассоциаций кукурузы. Наука 325, 737–740. doi: 10.1126/science.1174320
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Менг Х., Мужинский М.Г., Данилевская О.Н. (2011). FT-подобный ген ZCN8 действует как цветочный активатор и участвует в чувствительности к фотопериоду у кукурузы. Растительная клетка 23, 942–960. doi: 10.1105/tpc.110.081406
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Олсен, А. Н., Эрнст, Х. А., Леггио, Л. Л., и Скривер, К. (2005). Факторы транскрипции NAC: структурно различны, функционально разнообразны. Trends Plant Sci. 10, 79–87. doi: 10.1016/j.tplants.2004.12.010
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Поройко В., Spollen, W.G., Hejlek, L.G., Hernandez, A.G., LeNoble, M.E., Davis, G., et al. (2007). Сравнение региональных профилей транскриптов первичных корней кукурузы в условиях обильного полива и низкого водного потенциала. Дж. Экспл. Бот. 58, 279–289. doi: 10.1093/jxb/erl119
(2007). Сравнение региональных профилей транскриптов первичных корней кукурузы в условиях обильного полива и низкого водного потенциала. Дж. Экспл. Бот. 58, 279–289. doi: 10.1093/jxb/erl119
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Prasch, C.M., Ott, K.V., Bauer, H., Ache, P., Hedrich, R., and Sonnewald, U. (2015). Мутантные по ß-амилазе 1 растения арабидопсиса демонстрируют улучшенную засухоустойчивость из-за снижения распада крахмала в замыкающих клетках. Дж. Экспл. Бот. 66, 6059–6067. дои: 10.1093/jxb/erv323
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Рид, Дж. В., Нагатани, А., Элич, Т. Д., Фаган, М., и Чори, Дж. (1994). Фитохром А и фитохром В выполняют перекрывающиеся, но разные функции в развитии арабидопсиса. Физиол растений. 104, 1139–1149. doi: 10.1104/стр.104.4.1139
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Рейес, Дж. К., Муро-Пастор, М.I. и Флоренсио, Ф.Дж. (2004). Семейство транскрипционных факторов GATA у арабидопсиса и риса. Физиол растений. 134, 1718–1732. doi: 10.1104/стр.103.037788
К., Муро-Пастор, М.I. и Флоренсио, Ф.Дж. (2004). Семейство транскрипционных факторов GATA у арабидопсиса и риса. Физиол растений. 134, 1718–1732. doi: 10.1104/стр.103.037788
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Сари-Горла М., Краевски П., Ди Фонзо Н., Вилла М. и Фрова К. (1999). Генетический анализ засухоустойчивости кукурузы по молекулярным маркерам. II. Высота растений и цветение. Теор. заявл. Жене. 99, 289–295. дои: 10.1007/s001220051234
Полнотекстовая перекрестная ссылка | Академия Google
Шнабле, П.С., Уэр Д., Фултон Р.С., Штейн Дж.К., Вей Ф., Пастернак С. и соавт. (2009). Геном кукурузы B73: сложность, разнообразие и динамика. Наука 326, 1112–1115. doi: 10.1126/science.1178534
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Сеттер, Т.Л., Ян, Дж., Уорбертон, М., Рибо, Дж.М., Сюй, Ю., Сокинс, М., и др. (2010). Картирование генетической ассоциации идентифицирует полиморфизмы отдельных нуклеотидов в генах, которые влияют на уровни абсцизовой кислоты в цветочных тканях кукурузы во время засухи. Дж. Экспл. Бот. 62, 701–716. дои: 10.1093/jxb/erq308
Реферат PubMed | Полный текст перекрестной ссылки
Шанкар, Р., Бхаттачарджи, А., и Джейн, М. (2016). Анализ транскриптома различных сортов риса дает новое представление о реакциях на высыхание и засоление. науч. Респ. 6:23719. дои: 10.1038/srep23719
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Зоненберг, Н., и Хиннебуш, А. Г. (2009). Регуляция инициации трансляции у эукариот: механизмы и биологические мишени. Сотовый 136, 731–745. doi: 10.1016/j.cell.2009.01.042
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Стайгер, Д., и Браун, Дж. В. (2013). Альтернативный сплайсинг на пересечении биологического времени, развития и реакции на стресс. Растительная клетка 25, 3640–3656. doi: 10.1105/tpc.113.113803
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Су, З., Ма, X. , Го, Х., Сукиран, Н.Л., Го, Б., Ассманн, С.М., и другие. (2013). Развитие цветков в условиях засушливого стресса: морфологический и транскриптомный анализы показывают острые реакции и длительную акклиматизацию у арабидопсиса. Растительная клетка 25, 3785–3807. doi: 10.1105/tpc.113.115428
, Го, Х., Сукиран, Н.Л., Го, Б., Ассманн, С.М., и другие. (2013). Развитие цветков в условиях засушливого стресса: морфологический и транскриптомный анализы показывают острые реакции и длительную акклиматизацию у арабидопсиса. Растительная клетка 25, 3785–3807. doi: 10.1105/tpc.113.115428
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Тэтчер, С.Р., Данилевская, О.Н., Мэн, X., Битти, М., Застроу-Хейс, Г., Харрис, К., и др. (2016). Полногеномный анализ альтернативного сплайсинга во время развития и стресса от засухи у кукурузы. Физиол растений. 170, 586–599. doi: 10.1104/pp.15.01267
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Тирунавуккарасу Н., Хоссейн Ф., Арора К., Шарма Р., Ширига К., Миттал С. и др. (2014). Функциональные механизмы засухоустойчивости субтропической кукурузы ( Zea mays L.), выявленные с помощью полногеномного ассоциативного картирования. BMC Genomics 15:1182. дои: 10.1186/1471-2164-15-1182
дои: 10.1186/1471-2164-15-1182
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Трапнелл, К., Hendrickson, D.G., Sauvageau, M., Goff, L., Rinn, J.L., and Pachter, L. (2012). Дифференциальный анализ регуляции генов при разрешении транскриптов с помощью RNA-seq. Нац. Биотехнолог. 31, 46–53. doi: 10.1038/nbt.2450
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Урано К., Маруяма К., Огата Ю., Моришита Ю., Такеда М., Сакураи Н. и др. (2009). Характеристика регулируемых АБК глобальных реакций на обезвоживание у арабидопсиса с помощью метаболомики. Завод J. 57, 1065–1078. doi: 10.1111/j.1365-313X.2008.03748.x
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Виттинг-Сируп, К., Порс, Б.Т., Санделин, А., и Вааге, Дж. (2014). spliceR: пакет R для классификации альтернативного сплайсинга и прогнозирования потенциала кодирования на основе данных RNA-seq. БМС Биоинформ. 15:81. дои: 10.1186/1471-2105-15-81
15:81. дои: 10.1186/1471-2105-15-81
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Вахтер, А., Рюль, К., и Штауффер, Э. (2012). Роль белков, связывающих полипиримидиновый тракт, и других белков hnRNP в регуляции сплайсинга растений. Фронт. Растениевод. 3:81. doi: 10.3389/fpls.2012.00081
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Вайнгартнер, М., Суберт, К., и Зауэр, Н. (2011). LATE, белок цинковых пальцев C2h3, который действует как цветочный репрессор. Завод J. 68, 681–692. doi: 10.1111/j.1365-313X.2011.04717.x
Полнотекстовая перекрестная ссылка | Академия Google
Венг, X., Wang, L., Wang, J., Hu, Y., Du, H., Xu, C., et al. (2014). Количество зерен, высота растений и дата колошения7 являются центральными регуляторами роста, развития и реакции на стресс. Физиол растений. 164, 735–747. doi: 10.1104/стр.113.231308
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Wenkel, S. , Turck, F., Singer, K., Gissot, L., Le Gourrierec, J., Samach, A., et al. (2006). CONSTANS и комплекс, связывающий бокс CCAAT, имеют общий функционально важный домен и взаимодействуют, регулируя цветение арабидопсиса. Plant Cell Online 18, 2971–2984. doi: 10.1105/tpc.106.043299
, Turck, F., Singer, K., Gissot, L., Le Gourrierec, J., Samach, A., et al. (2006). CONSTANS и комплекс, связывающий бокс CCAAT, имеют общий функционально важный домен и взаимодействуют, регулируя цветение арабидопсиса. Plant Cell Online 18, 2971–2984. doi: 10.1105/tpc.106.043299
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Xu, J., Yuan, Y., Xu, Y., Zhang, G., Guo, X., Wu, F., et al. (2014). Идентификация генов-кандидатов засухоустойчивости путем ресеквенирования всего генома кукурузы. BMC Растение Биол. 14:83. дои: 10.1186/1471-2229-14-83
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Сюэ, В., Син, Ю., Вэн, X., Чжао Ю., Тан В., Ван Л. и др. (2008). Естественная изменчивость Ghd7 является важным регулятором срока колошения и потенциальной урожайности риса. Нац. Жене. 40, 761–767. doi: 10.1038/ng.143
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Ян, К. , Лю, П., Ву, К., Ян, Г., Сюй, Р., Го, К., и др. (2012). Индуцированный стрессом альтернативный сплайсинг обеспечивает механизм регуляции процессинга микроРНК в Arabidopsis thaliana . мол.Клетка. 48, 521–531. doi: 10.1016/j.molcel.2012.08.032
, Лю, П., Ву, К., Ян, Г., Сюй, Р., Го, К., и др. (2012). Индуцированный стрессом альтернативный сплайсинг обеспечивает механизм регуляции процессинга микроРНК в Arabidopsis thaliana . мол.Клетка. 48, 521–531. doi: 10.1016/j.molcel.2012.08.032
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Ю, С.Ю., Ким, Ю., Ким, С.Ю., Ли, Дж.С., и Ан, Дж.Х. (2007). Контроль времени цветения и реакции на холод с помощью белка NAC-домена у арабидопсиса. PLoS ONE 2:e642. doi: 10.1371/journal.pone.0000642
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Ю, Дж., Холланд, Дж. Б., МакМаллен, М. Д., и Баклер, Э.С. (2008). Генетический дизайн и статистическая мощность картирования вложенных ассоциаций у кукурузы. Генетика 178, 539–551. doi: 10.1534/genetics.107.074245
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Юэ, Г., Чжуан, Ю., Ли, З., Сунь, Л., и Чжан, Дж. (2008). Дифференциальный анализ экспрессии генов листьев кукурузы на стадии колошения в ответ на дефицит воды. Бионауч. 28, 125–134. дои: 10.1042/BSR20070023
(2008). Дифференциальный анализ экспрессии генов листьев кукурузы на стадии колошения в ответ на дефицит воды. Бионауч. 28, 125–134. дои: 10.1042/BSR20070023
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Чжэн, Дж., Zhao, J., Tao, Y., Wang, J., Liu, Y., Fu, J., et al. (2004). Выделение и анализ генов, индуцированных водным стрессом, в проростках кукурузы с помощью субтрактивной ПЦР и макрочипа кДНК. Завод Мол. биол. 55, 807–823. doi: 10.1007/s11103-005-1969-9
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Zhuang, Y., Ren, G., Yue, G., Li, Z., Qu, X., Hou, G., et al. (2007). Влияние стресса, вызванного дефицитом воды, на транскриптомы развивающихся незрелых початков и метелок кукурузы. Представитель клеток растений 26, 2137–2147. doi: 10.1007/s00299-007-0419-3
Реферат PubMed | Полный текст перекрестной ссылки | Академия Google
Зийомо, К., и Бернардо, Р. (2013). Засухоустойчивость кукурузы: непрямой отбор по вторичным признакам против полногеномного отбора. Растениеводство. 53, 1269. doi: 10.2135/cropsci2012.11.0651
Засухоустойчивость кукурузы: непрямой отбор по вторичным признакам против полногеномного отбора. Растениеводство. 53, 1269. doi: 10.2135/cropsci2012.11.0651
Полнотекстовая перекрестная ссылка | Академия Google
Цветение и детерминантность кукурузы | Журнал экспериментальной ботаники
Аннотация
Все органы растений образуются из меристем, групп стволовых клеток, расположенных на кончиках корней и побегов.Неопределенные меристемы образуют неопределенное количество органов, тогда как детерминированные меристемы потребляются после образования определенного числа органов. Кукуруза является идеальной системой для изучения генетического контроля за судьбой меристем из-за вклада детерминантных и индетерминантных меристем в общее соцветие. Здесь представлен обзор последних работ по поддержанию меристем и спецификации органов кукурузы. Генетические сети, такие как компоненты CLAVATA поддержания меристемы и программа развития цветка ABC, сохраняются между травами и эвдикотами. Кукуруза и рис, по-видимому, имеют законсервированные механизмы поддержания меристемы и идентичности органов. Другие пути, такие как детерминация пола, скорее всего, обнаружены только у кукурузы с отдельными мужскими и женскими цветками. Богатая генетическая история привела к созданию большой коллекции мутантов кукурузы. Появление геномных инструментов и синтении у трав теперь позволяет изолировать гены, лежащие в основе архитектуры соцветия, и дает возможность сравнивать функции у покрытосеменных растений.
Кукуруза и рис, по-видимому, имеют законсервированные механизмы поддержания меристемы и идентичности органов. Другие пути, такие как детерминация пола, скорее всего, обнаружены только у кукурузы с отдельными мужскими и женскими цветками. Богатая генетическая история привела к созданию большой коллекции мутантов кукурузы. Появление геномных инструментов и синтении у трав теперь позволяет изолировать гены, лежащие в основе архитектуры соцветия, и дает возможность сравнивать функции у покрытосеменных растений.
Введение
Замечательным аспектом растений является их способность производить новые органы на протяжении всей жизни.Эта способность достигается действием меристем, групп самообновляющихся стволовых клеток, расположенных на верхушках побегов и корней (Steeves and Sussex, 1989). Деление меристемы дает начало клеткам с разной судьбой. Клетки в центре меристемы, центральной зоне, продолжают пополнять меристему, сохраняя определенный размер. Клетки на периферии меристемы находятся в морфогенетической зоне, из которой со временем возникают органы. Равновесие этих двух процессов, органогенеза и самовоспроизводства, гарантирует пролонгированную активность, и такая меристема называется индетерминантной.Напротив, детерминированные меристемы, такие как те, которые производят цветы, потребляются после образования определенного количества органов.
Клетки на периферии меристемы находятся в морфогенетической зоне, из которой со временем возникают органы. Равновесие этих двух процессов, органогенеза и самовоспроизводства, гарантирует пролонгированную активность, и такая меристема называется индетерминантной.Напротив, детерминированные меристемы, такие как те, которые производят цветы, потребляются после образования определенного количества органов.
Соцветие кукурузы представляет собой прекрасную модель для изучения контроля развития меристем, поскольку оно сформировано как индетерминантными, так и детерминантными меристемами. Кроме того, кукуруза имеет богатую генетическую историю, и было описано несколько мутаций, влияющих на отдельные стадии развития соцветия (Neuffer et al. , 1997). Эти мутанты могут иметь аномальный размер меристемы или неверную спецификацию идентичности органа, или и то, и другое.Генетика развития соцветий и цветков у кукурузы и других злаков недавно была рассмотрена другими авторами (McSteen et al. , 2000; Bommert et al. , 2005 a ). Здесь кратко изложены современные знания о развитии соцветий травы и сделан обзор последних работ. Основное внимание уделяется кукурузе, но также включены некоторые открытия в области риса.
, 2000; Bommert et al. , 2005 a ). Здесь кратко изложены современные знания о развитии соцветий травы и сделан обзор последних работ. Основное внимание уделяется кукурузе, но также включены некоторые открытия в области риса.
Основной единицей архитектуры соцветия травы является колосок, компактная пазушная ветвь, состоящая из двух прицветников, на которых расположены от одного до нескольких редуцированных цветков (Clifford, 1987).Кукуруза является однодомным растением, которое производит мужские цветки на концевой метелке (рис. 1А) и женские цветки на боковых початках (рис. 1Б), которые возникают в пазухах вегетативных листьев. Кисточка дает начало нескольким длинным неопределенным ветвям у основания, в то время как початок состоит из одного шипа без длинных ветвей. Главный шип и ветви кисточки, а также весь колос дают короткие ветви (пары колосков), несущие два колоска (рис. 1C, D, 2A–D). Ветви и пары колосков возникают в пазухах маленьких неразвитых листьев, называемых прицветниками. У кукурузы колоски и парные меристемы колосков считаются детерминированными, поскольку они образуют определенное количество органов (Vollbrecht et al. , 2005).
У кукурузы колоски и парные меристемы колосков считаются детерминированными, поскольку они образуют определенное количество органов (Vollbrecht et al. , 2005).
Рис. 1.
Соцветия кукурузы дикого типа. (A) Кисточка с парами колосков (вставка) на ветвях и главном колосе. (B) Ухо, показывающее ряды ядер. (C) Сканирующая электронная микроскопия зачатка кисточки, инициирующего меристемы пары колосков. (D) Сканирующая электронная микроскопия уха. Spm: парная меристема колоска.См: колосковая меристема.
Рис. 1.
Соцветия кукурузы дикого типа. (A) Кисточка с парами колосков (вставка) на ветвях и главном колосе. (B) Ухо, показывающее ряды ядер. (C) Сканирующая электронная микроскопия зачатка кисточки, инициирующего меристемы пары колосков. (D) Сканирующая электронная микроскопия уха. Spm: парная меристема колоска. См: колосковая меристема.
Размер меристем соцветий и уход за ними
В отличие от своего предка teosinte, цветение которого вызвано коротким днем, кукуруза переходит от вегетативной к репродуктивной фазе после образования фиксированного количества листьев (Irish and Nelson, 1988). Наиболее важным известным регулятором перехода к репродуктивной стадии у кукурузы является индетерминантный1 ( id1 ). Ген id1 кодирует белок цинковых пальцев, который вырабатывается в молодых листьях. ID1 функционирует неавтономно, чтобы подать сигнал апикальной меристеме побега (SAM) для перехода к репродуктивной стадии. Мутанты id1 образуют гораздо больше листьев, чем кукуруза дикого типа, и демонстрируют вегетативную реверсию в метелке с проростками, возникающими из мужских колосков, часто с корнями (Colasanti et al., 1998).
Наиболее важным известным регулятором перехода к репродуктивной стадии у кукурузы является индетерминантный1 ( id1 ). Ген id1 кодирует белок цинковых пальцев, который вырабатывается в молодых листьях. ID1 функционирует неавтономно, чтобы подать сигнал апикальной меристеме побега (SAM) для перехода к репродуктивной стадии. Мутанты id1 образуют гораздо больше листьев, чем кукуруза дикого типа, и демонстрируют вегетативную реверсию в метелке с проростками, возникающими из мужских колосков, часто с корнями (Colasanti et al., 1998).
в Eudicots Antirrhinum и Arabidopsiss , гены Floricaula ( FLO ) и Leasty ( LFY ) необходимы для производства цветов (Coen et al. , 1990; Weigel и др. , 1992). Мутанты flo производят ветви соцветия в положении цветков, в то время как мутанты lfy производят цветки с чертами соцветия. Кукуруза имеет дубликаты ортологов FLO/LFY , zea flo/lfy1 ( zfl1 ) и zfl2 . Фенотипический анализ мутантов zfl1 и zfl2 показывает, что их функция несколько консервативна у кукурузы. В то время как одиночные мутанты zfl1 и zfl2 имеют мягкие фенотипы, двойные мутанты не претерпевают нормального перехода к репродуктивному развитию и имеют аномальные терминальные соцветия. Их называют «уши с кисточками», потому что они представляют собой разветвленные соцветия с женскими ветвями, покрытыми листьями шелухи, которые заканчиваются колосом мужских цветков.Развитие колосков, цветков и цветковых чешуй у мутантов zfl1 / zfl2 выглядит нормальным, но в цветках обнаруживаются различные дефекты, связанные с отсутствием детерминированности и органной идентичности: нормальные плодолистики не формируются, вместо них формируется женская меристема цветка. производит несколько «карпеллоидных» органов с шелками и вегетативными выростами. Точно так же в метелке цветовые чешуи и цветовые чешуи нормальные, но тычинки не развиваются и иногда заменяются цветковыми чешуями или похожими на цветковые чешуи органами (Bomblies et al.
Фенотипический анализ мутантов zfl1 и zfl2 показывает, что их функция несколько консервативна у кукурузы. В то время как одиночные мутанты zfl1 и zfl2 имеют мягкие фенотипы, двойные мутанты не претерпевают нормального перехода к репродуктивному развитию и имеют аномальные терминальные соцветия. Их называют «уши с кисточками», потому что они представляют собой разветвленные соцветия с женскими ветвями, покрытыми листьями шелухи, которые заканчиваются колосом мужских цветков.Развитие колосков, цветков и цветковых чешуй у мутантов zfl1 / zfl2 выглядит нормальным, но в цветках обнаруживаются различные дефекты, связанные с отсутствием детерминированности и органной идентичности: нормальные плодолистики не формируются, вместо них формируется женская меристема цветка. производит несколько «карпеллоидных» органов с шелками и вегетативными выростами. Точно так же в метелке цветовые чешуи и цветовые чешуи нормальные, но тычинки не развиваются и иногда заменяются цветковыми чешуями или похожими на цветковые чешуи органами (Bomblies et al. , 2003).
, 2003).
Одним из ключевых процессов поддержания меристемы является путь CLAVATA (CLV), первоначально описанный у Arabidopsis и названный в честь трех генов: CLV1 , CLV2 и CLV3 . Мутации в этих генах вызывают увеличение меристем побегов и цветков, что приводит к появлению цветков с дополнительными частями цветков (Clark и др. , 1993, Clark и др. , 1995, Kayes and Clark, 1998). CLV1 и C LV2 принадлежат к большому семейству генов и оба содержат трансмембранный домен. CLV1 представляет собой рецептороподобную киназу (RLK) с богатой лейцином повторяющейся областью (LRR), а CLV2 представляет собой рецептороподобный белок (RLP) с LRR, но, в отличие от CLV1 , у него отсутствует цитоплазматическая хвост и не имеет киназной функции (Jeong et al. , 1999). CLV3 кодирует внеклеточный белок, который предположительно взаимодействует с CLV1 и CLV2 с образованием комплекса, запускающего сигнальный путь в центральной зоне SAM, что приводит к ограничению накопления стволовых клеток через фактор транскрипции WUSCHEL ( Карлес и Флетчер, 2003).
В последние годы стало очевидным, что большая часть пути CLV сохраняется между Arabidopsis и травами. У кукурузы ген толстая кисточка dwarf1 кодирует LRR-RLK и является предполагаемым ортологом CLV1 (Bommert et al. , 2005 b ). Другой ген кукурузы, фасциированного колоса 2 , кодирует LRR-RLP, сходный с CLV2 (Taguchi-Shiobara et al. , 2001). Фенотипы соцветий мутантов td1 и fea2 сходны.Мутации приводят к фасциальным початкам, увеличению плотности колосков в метелке за счет более толстого стержня и увеличению числа тычинок в мужских цветках. Початки имеют увеличенное количество семенных рядов, они короче и толще, чем у дикого типа. Эти фенотипы согласуются с наблюдаемым увеличением размеров всех меристем соцветий мутантов td1 и fea2 . Доминантный мутант кукурузы, вызывающий фасциацию початка, Fasciated ear1 ( Fas1 ) (рис.3F), недавно был идентифицирован, и его роль в пути CLV изучается (China Lunde, неопубликованные результаты).
Мутации в ортологах риса CLV1 и CLV3 обнаруживают сходные фенотипы с мутантами кукурузы td1 и fea2 , особенно увеличение апикальных и цветочных меристем. Ген FLORAL ORGAN NUMBER1 ( FON1 ), который кодирует LRR-RLK, сходный с CLV1 и td1 , экспрессируется в вегетативных и репродуктивных меристемах и во всех зачатках органов цветка (Suzaki et al., 2004). Несмотря на широкий характер экспрессии, мутации в FON1 влияют только на меристемы цветков за счет увеличения числа лепестков, цветковых чешуй, тычинок и пестиков. Мутации в FON4 , гене с последовательностью, сходной с CLV3 , вызывают увеличение апикальных побегов, соцветий и цветочных меристем (Chu et al. , 2006). Следовательно, мутанты FON4 имеют более толстые стебли, дополнительные ветви соцветия и экстрафлоральные органы. FON4 экспрессируется в центральной зоне вегетативных, соцветий и цветочных меристем, подобно CLV3 . Другой рис CLV3 -подобный ген, FON2, , имеет широкий паттерн экспрессии, но его потеря функции влияет только на спецификацию числа органов цветка, в то время как соцветия являются нормальными (Suzaki et al. , 2004). Это говорит о том, что в вегетативных меристемах и меристемах соцветий FON4 действует избыточно с FON2, ограничивая размер меристемы, но оба необходимы в цветочных меристемах для правильного формирования цветка.
Другой рис CLV3 -подобный ген, FON2, , имеет широкий паттерн экспрессии, но его потеря функции влияет только на спецификацию числа органов цветка, в то время как соцветия являются нормальными (Suzaki et al. , 2004). Это говорит о том, что в вегетативных меристемах и меристемах соцветий FON4 действует избыточно с FON2, ограничивая размер меристемы, но оба необходимы в цветочных меристемах для правильного формирования цветка.
Противоположная группа мутантов кукурузы не дает ветвей и колосков. бесплодное соцветие2 ( bif2 ) мутанты практически не имеют ветвей как в колосе, так и в метелках. Прицветники не затрагиваются, что продемонстрировано двойным мутантом tasselsheath , который дает увеличенные прицветники (McSteen and Hake, 2001). bif2 кодирует серин/треониновую протеинкиназу, коортологичную PID из Arabidopsis (P McSteen and S Hake, неопубликованные данные). PID отвечает за полярную локализацию белков PIN-носителей оттока ауксина (Friml et al. , 2004), и оба мутанта PIN и PID с потерей функции имеют соцветия без листьев, пазушных ветвей и цветков (Galweiler et al. , 1998; Christensen et al. , 2000). Мутанты kn1 с потерей функции проявляют умеренный бесплодный фенотип. Уши часто не формируются, а если и формируются, то имеют мало зёрен и дополнительных шёлков. На кисточках меньше ветвей и пар колосков (Kerstetter et al. , 1997).
, 2004), и оба мутанта PIN и PID с потерей функции имеют соцветия без листьев, пазушных ветвей и цветков (Galweiler et al. , 1998; Christensen et al. , 2000). Мутанты kn1 с потерей функции проявляют умеренный бесплодный фенотип. Уши часто не формируются, а если и формируются, то имеют мало зёрен и дополнительных шёлков. На кисточках меньше ветвей и пар колосков (Kerstetter et al. , 1997).
Мутанты гена риса LAX PANICLE ( LAX ) и его ортолога кукурузы barren stalk1 ( ba1 ) также демонстрируют снижение образования пазушных ветвей.Хотя дефекты соцветий схожи, влияние на вегетативное ветвление различно. Мутантные растения ba1 не продуцируют никаких побегов, в то время как кустистость у мутантов lax затрагивается лишь слабо (Komatsu et al. , 2003; a ; Gallavotti et al. , 2004). Это различие, вероятно, связано с избыточностью LAX и SMALL PANICLE ( SPA ), поскольку двойные мутанты lax/spa имеют значительно меньшее количество побегов (Komatsu et al. , 2003 и ).
, 2003 и ).
Паттерн экспрессии ra2 , маркера очень ранних стадий пазушного ветвления (Bortiri et al. , 2006), можно использовать для изучения стадии, на которой мутант отклоняется от нормального развития. Экспрессия ra2 в метелках bif2 показывает, что этот мутант инициирует меньше, чем нормальные меристемы, но они имеют нормальный размер. Следовательно, bif2 соцветий образуют несколько «побеговых» пазушных ветвей и пар колосков (McSteen, Hake, 2001).С другой стороны, метелки ba1 имеют нормальное распределение пазушных меристем, определяемое доменами экспрессии ra2 , но размер зачатка меристемы меньше, что указывает на то, что меристемы мутантов ba1 не могут расти, потому что они не достигают критического размера (E Bortiri et al. , неопубликованные результаты).
Инициация и детерминация пазушной меристемы
Пары колосков являются производным признаком, присутствующим только в трибе Andropogoneae, монофилетической группе, в которую входят такие виды, как кукуруза, сорго и сахарный тростник (LeRoux and Kellogg, 1999; Group, 2001). И пары колосков, и индетерминантные ветви происходят от пазушных меристем; однако, в отличие от других пазушных меристем кистевидных соцветий, меристемы пар колосков заканчиваются после образования двух колосков. По этой причине они считаются детерминированными. Филогенетическое расположение пар колосков предполагает, что у Andropogoneae возникла новая генетическая программа, определяющая судьбу детерминированных пазушных меристем.
И пары колосков, и индетерминантные ветви происходят от пазушных меристем; однако, в отличие от других пазушных меристем кистевидных соцветий, меристемы пар колосков заканчиваются после образования двух колосков. По этой причине они считаются детерминированными. Филогенетическое расположение пар колосков предполагает, что у Andropogoneae возникла новая генетическая программа, определяющая судьбу детерминированных пазушных меристем.
Мутанты ramosa ( ra1 , ra2 , ra3 ) обеспечивают исходный материал для изучения молекулярной основы программы развития пар колосков.Все мутанты ra имеют пазушные меристемы с повышенной индетерминантностью. Початки сильных мутантных аллелей ra образуют ветви, а в метелках пары колосков заменяются индетерминантными ветвями. Длинные ветви у основания кисточки демонстрируют повышенную степень ветвления (рис. 3А–С).
ra1 кодирует Cys2-His2 цинковый палец специфичного для растений подкласса EPF и экспрессируется в основании меристемы пары колосков (Vollbrecht et al. , 2005). ra2 кодирует специфический для растений фактор транскрипции домена LOB, содержащий область, богатую цистеином, и область, подобную лейциновой молнии. RA2 кукурузы, сорго, ячменя и риса имеет консервативный С-концевой домен в дополнение к домену LOB (Bortiri et al. , 2006). ra2 экспрессируется в зачатках неопределенных длинных ветвей, меристемах пар колосков и меристемах колосков. ra3 кодирует трегалозо-6-фосфатфосфатазу (TPP) и экспрессируется в основании меристем пары колосков (Satoh et al., 2006). Уровни герметики RA1 очень низкие в RA2 и RA3 мутантные соцветия и RA1 / RA2 , и RA1 / RA1 Двойные мутаты RA3 показывают аддитивный фенотип (Vollbrecht et al. , 2005; Satoh и др. , 2006). Кроме того, RA2 экспрессия является нормальным в RA1 и RA3 мутантов (Bortiri et al. , 2006) и Уровни транскрипта RA3 не изменяются в RA2 и RA1 Muttants (SATOH и другие. , 2006). Эти данные предполагают, что ra2 и ra3 находятся выше ra1 , но по разным путям, и оба необходимы для нормальной транскрипции ra1 . Это открытие может объяснить, почему ra1 не экспрессируется в меристемах ветвей, которые имеют ra2 , но не экспрессируют ra3 . Ортологи ra2 и ra3 были обнаружены в других злаках, в том числе в рисе, и они имеют схожий характер экспрессии в этих видах злаков (Bortiri et al., 2006 г.; Сато и др. , 2006). Однако ортолог ra1 не был обнаружен в рисе или других травах за пределами Andropogoneae (E. Vollbrecht, EA Kellogg и S. Malcomber, неопубликованные результаты). Судя по консервативности их последовательностей и паттернов экспрессии, кажется, что RA2 и RA3 были рекрутированы для регуляции ra1 , гена, функция которого возникла у Andropogoneae, и все три действуют вместе, чтобы навязать детерминированную судьбу меристемам пар колосков.
, 2005). ra2 кодирует специфический для растений фактор транскрипции домена LOB, содержащий область, богатую цистеином, и область, подобную лейциновой молнии. RA2 кукурузы, сорго, ячменя и риса имеет консервативный С-концевой домен в дополнение к домену LOB (Bortiri et al. , 2006). ra2 экспрессируется в зачатках неопределенных длинных ветвей, меристемах пар колосков и меристемах колосков. ra3 кодирует трегалозо-6-фосфатфосфатазу (TPP) и экспрессируется в основании меристем пары колосков (Satoh et al., 2006). Уровни герметики RA1 очень низкие в RA2 и RA3 мутантные соцветия и RA1 / RA2 , и RA1 / RA1 Двойные мутаты RA3 показывают аддитивный фенотип (Vollbrecht et al. , 2005; Satoh и др. , 2006). Кроме того, RA2 экспрессия является нормальным в RA1 и RA3 мутантов (Bortiri et al. , 2006) и Уровни транскрипта RA3 не изменяются в RA2 и RA1 Muttants (SATOH и другие. , 2006). Эти данные предполагают, что ra2 и ra3 находятся выше ra1 , но по разным путям, и оба необходимы для нормальной транскрипции ra1 . Это открытие может объяснить, почему ra1 не экспрессируется в меристемах ветвей, которые имеют ra2 , но не экспрессируют ra3 . Ортологи ra2 и ra3 были обнаружены в других злаках, в том числе в рисе, и они имеют схожий характер экспрессии в этих видах злаков (Bortiri et al., 2006 г.; Сато и др. , 2006). Однако ортолог ra1 не был обнаружен в рисе или других травах за пределами Andropogoneae (E. Vollbrecht, EA Kellogg и S. Malcomber, неопубликованные результаты). Судя по консервативности их последовательностей и паттернов экспрессии, кажется, что RA2 и RA3 были рекрутированы для регуляции ra1 , гена, функция которого возникла у Andropogoneae, и все три действуют вместе, чтобы навязать детерминированную судьбу меристемам пар колосков. Функция ra2 и ra3 в других травах еще не известна, но предполагается, что они модулируют степень ветвления.
Функция ra2 и ra3 в других травах еще не известна, но предполагается, что они модулируют степень ветвления.
Другими мутантами с потерей детерминантности как в метелках, так и в колосе являются разветвленный шелковистый1 (bd1 ) , индетерминантный колосок1 (ids1 ) , метелочное семя4 (ts4 ), пушистосемянный 7 мутант Tassel 6 доминантный (Ц6 ). bd1 и ids1 кодируют белки, содержащие один или два домена АР2 соответственно, и оба влияют на меристему колоска (Chuck et al., 1998, 2002), однако судьбы этих меристем различаются. Мутанты bd1 обнаруживают очень яркий фенотип колоса, в котором колосок заменяется стерильной индетерминантной ветвью. Каждая ветвь образует парные меристемы колоска, подобные ветвям кисточки. В метелке меристемы колосков также недетерминированы, но образуют колоски с двудольным рисунком, что в конечном итоге дает фертильные цветки (Chuck et al. , 2002). Мутант риса вьющаяся метелка является ортологом bd1 со сходным мутантным фенотипом (Komatsu et al., 2003 б ). Мутанты ids1 также имеют потерю детерминантности в СМ, однако СМ сохраняет свою идентичность и образует дополнительные цветки и колосковые чешуи. Ухо и кисточка похожи, хотя початок не оплодотворяемый (Chuck et al. , 1998). Мутант fuzzy tassel также образует дополнительные цветки на колосок и множественные стерильные части цветка, но, в отличие от ids1 , у него отсутствуют нормальные чешуйки (Fig. 3F).
Мутант риса вьющаяся метелка является ортологом bd1 со сходным мутантным фенотипом (Komatsu et al., 2003 б ). Мутанты ids1 также имеют потерю детерминантности в СМ, однако СМ сохраняет свою идентичность и образует дополнительные цветки и колосковые чешуи. Ухо и кисточка похожи, хотя початок не оплодотворяемый (Chuck et al. , 1998). Мутант fuzzy tassel также образует дополнительные цветки на колосок и множественные стерильные части цветка, но, в отличие от ids1 , у него отсутствуют нормальные чешуйки (Fig. 3F).
bd1 экспрессируется только в колосе и метелках в очень узком участке на боку меристемы колоска (Chuck et al., 2002). Экспрессия bd1 наблюдается, когда меристема пары колосков делится с образованием меристемы колоска. Выражение появляется сначала у основания меристемы одного колоска, затем у основания другого. Выражение отмечает положение между колосковой чешуей и меристемой колоска. ids1 экспрессируется более широко, хотя и имеет узкую область действия. Она выражена в SPM и SM, а также в лепестках, цветковых чешуях и инициалях тычинок. Экспрессия не наблюдается в плодолистиках и чешуях (Chuck et al., 1998).
ids1 экспрессируется более широко, хотя и имеет узкую область действия. Она выражена в SPM и SM, а также в лепестках, цветковых чешуях и инициалях тычинок. Экспрессия не наблюдается в плодолистиках и чешуях (Chuck et al., 1998).
Ц6 и ц4 имеют сходные фенотипы (рис. 3Г, Д). Они оба имеют феминизированные кисточки (см. ниже обсуждение определения пола), но могут иметь мужскую фертильность в зависимости от врожденного происхождения (Э. Бортири и С. Хейк, личные наблюдения). У этих двух мутантов детерминированность меристем нарушена на разных стадиях развития соцветия. Анализ Erin Irish показывает, что у ts4 парных меристем колосков трансформируются в индетерминантные ветви, несущие дополнительные пары колосков, а у Ts6 меристемы колосков на ножке дают больше цветков (Irish, 1997).SEM-анализ предполагает, что паттерны ветвления в ts4 не такие регулярные, как у мутантов ids1 или bd1 (Irish, 1997; G Chuck and S Hake, неопубликованные результаты).
Определение пола в цветках кукурузы
Все цветки кукурузы образуют белую лепестку, цветковую чешую, две лодыжки, три тычинки и три плодолистика, которые сливаются в один пестик. После инициации зачатки пестика в цветках-метелках прерываются, а зачатки тычинок в колосе обнаруживают остановку клеточного цикла (Dellaporta and Calderon-Urrea, 1994).Кроме того, нижний цветочек колоса также останавливается. В результате колоски-метелки несут два функциональных тычиночных цветка, а в колосе до зрелости развивается только один пестичный цветок.
Было обнаружено несколько мутантов, которые изменяют определение пола мужских или женских цветков. Особый класс карликовых растений — андромонодомные, с мужскими цветками в метелке и совершенными цветками в колосе. Большинство андромоноэциальных карликов дефектны в биосинтезе ГА (Phinney, 1956), хотя доминантный мутант D8 дефектен в реакции на ГА (Harberd and Freeling, 1989). D8 гомологичен GAI из Arabidopsis и содержит N-концевую делецию домена DELLA (Peng et al. , 1999). Понимание того, как ГК регулирует детерминацию пола, неизвестно, но обнаружение того, что уровни ГК в 100 раз ниже в развивающихся колосках кистей, предполагает, что низкие уровни ГК необходимы для развития тычиночных цветков, а более высокие уровни вызывают аборты тычинок (Rood and Pharis, 1980). ). Дополнительным эффектом мутантов с дефицитом GA у кукурузы является уменьшение количества ветвей метелок.Это наиболее очевидно в пыльниковом ухе 1, который кодирует ent-kaurene synthase, фермент, который катализирует раннюю стадию биосинтеза GA (Bensen et al. , 1995). Таким образом, ГК может регулировать не только аборты тычинок в колосе, но и ветвление метелок.
, 1999). Понимание того, как ГК регулирует детерминацию пола, неизвестно, но обнаружение того, что уровни ГК в 100 раз ниже в развивающихся колосках кистей, предполагает, что низкие уровни ГК необходимы для развития тычиночных цветков, а более высокие уровни вызывают аборты тычинок (Rood and Pharis, 1980). ). Дополнительным эффектом мутантов с дефицитом GA у кукурузы является уменьшение количества ветвей метелок.Это наиболее очевидно в пыльниковом ухе 1, который кодирует ent-kaurene synthase, фермент, который катализирует раннюю стадию биосинтеза GA (Bensen et al. , 1995). Таким образом, ГК может регулировать не только аборты тычинок в колосе, но и ветвление метелок.
tassel seed1 (ts1 ) и ts2 имеют полностью феминизированные кисточки независимо от фона. Кроме того, нижний цветочек початка не прерывается. ts2 кодирует короткоцепочечную алкогольдегидрогеназу, которая, как предполагается, приводит к выкидышу пестика (DeLong et al. , 1993). Экспрессия ts2 не наблюдается у мутантов ts1 , поэтому считается, что ts1 действует выше ts2 (Calderon-Urrea and Dellaporta, 1999). ts2 экспрессируется во всех зачатках пестика, включая зачатки початка, таким образом, Кальдерон-Урреа и Деллапорта предполагают, что верхний цветочек початка защищен от посттранскрипционной гибели клеток пестика, вызванной TS2 (Calderon-Urrea and Dellaporta, 1999). . silkless1 ( sk ) имеет фенотип, противоположный ts1 и ts2; кисточка нормальная, но колосья без пестиков и поэтому стерильны.Двойные мутанты с ts2 показывают, что ts2 является эпистатическим по отношению к sk (Jones, 1932; Veit et al. , 1991; Irish et al. , 1994; Calderon-Urrea, 1999). Эти результаты предполагают, что SK негативно регулирует TS2 в верхнем цветке початка, таким образом, только этот цветочек защищен от гибели клеток, опосредованной TS2.
, 1993). Экспрессия ts2 не наблюдается у мутантов ts1 , поэтому считается, что ts1 действует выше ts2 (Calderon-Urrea and Dellaporta, 1999). ts2 экспрессируется во всех зачатках пестика, включая зачатки початка, таким образом, Кальдерон-Урреа и Деллапорта предполагают, что верхний цветочек початка защищен от посттранскрипционной гибели клеток пестика, вызванной TS2 (Calderon-Urrea and Dellaporta, 1999). . silkless1 ( sk ) имеет фенотип, противоположный ts1 и ts2; кисточка нормальная, но колосья без пестиков и поэтому стерильны.Двойные мутанты с ts2 показывают, что ts2 является эпистатическим по отношению к sk (Jones, 1932; Veit et al. , 1991; Irish et al. , 1994; Calderon-Urrea, 1999). Эти результаты предполагают, что SK негативно регулирует TS2 в верхнем цветке початка, таким образом, только этот цветочек защищен от гибели клеток, опосредованной TS2. Двойные мутанты между ts2 и D8-Mp1 демонстрируют аддитивный фенотип с идеальными цветками (тычинками и пестиками) как в колосе, так и в метелке (Veit et al., 1991; Ирландский и др. , 1994). Эти результаты показывают, что пути TS2 и GA действуют независимо, и развитие пестика в метелке не зависит от потери тычинок.
Двойные мутанты между ts2 и D8-Mp1 демонстрируют аддитивный фенотип с идеальными цветками (тычинками и пестиками) как в колосе, так и в метелке (Veit et al., 1991; Ирландский и др. , 1994). Эти результаты показывают, что пути TS2 и GA действуют независимо, и развитие пестика в метелке не зависит от потери тычинок.
Спецификация органов цветков кукурузы
Модель развития цветка ABC была первоначально описана для эвдикотов Arabidopsis и Antirrhinum . Эта модель утверждает, что гены класса A определяют судьбу чашелистиков в первой мутовке цветка, гены A плюс B определяют лепестки во второй мутовке, гены B плюс C дают начало тычинкам, и только гены C необходимы для развития плодолистика в четвертой мутовке. (Коэн и Мейеровиц, 1991).Недавно эта модель была расширена за счет включения генов класса D, ответственных за развитие семязачатков, и генов класса Е, необходимых для нормальной экспрессии всех вышеупомянутых генов. За исключением гена A-класса APETALA2 , все эти гены являются членами семейства транскрипционных факторов MADS-box и действуют путем образования димеров и комплексов более высокого порядка (de Folter et al. , 2005). .
За исключением гена A-класса APETALA2 , все эти гены являются членами семейства транскрипционных факторов MADS-box и действуют путем образования димеров и комплексов более высокого порядка (de Folter et al. , 2005). .
silky1 ( si1 ) представляет собой ген MADS-box, связанный с Arabidopsis APETALA3 и Antirrhinum DEFICIENS (B-класс).Мутации в si1 превращают лодикулы в прицветниковые органы, напоминающие цветковые чешуи или цветковые цветковые чешуи (Ambrose et al. , 2000), а тычинки в пестики. Этот фенотип похож на потерю функции генов B-класса у эвдикотов. si1 экспрессируется в центре цветочной меристемы во время образования чешуи и цветка. Позже экспрессия ограничивается областью цветочной меристемы, которая дает начало лодикулам и тычинкам. Тот факт, что SILKY1 обладает биохимическими свойствами белков B-класса и может спасти мутант Arabidopsis ap3 , показывает, что функция B-класса сохраняется между травами и Arabidopsis (Whipple et al. , 2004). Гены C-класса
, 2004). Гены C-класса
AGAMOUS и PLENA , Arabidopsis и Antirrhinum определяют идентичность тычинок и плодолистиков в третьем и четвертом оборотах, а также придают детерминированность цветочной меристемы. У мутантов ag и ple цветки образуют чашелистики и лепестки повторяющимся образом (Yanofsky et al. , 1990; Bradley et al. , 1993). Кукуруза и рис имеют дублированные AG -подобные гены, и они, по-видимому, развили частичную субфункционализацию.Мутации в zag1 вызывают неопределенный рост зачатков пестика, дающий начало более чем одному шелку и недифференцированным массам ткани внутри яичника (Mena et al. , 1996). В метелке иногда развиваются некоторые шелка, указывающие на то, что аборт пестика не завершен, но тычинки нормальные. Отсутствие влияния на идентичность тычинок было объяснено присутствием zmm2 , другого бокс-гена MADS, очень похожего на AG и PLE . Паттерны экспрессии zag1 и zmm2 в значительной степени не перекрываются, поскольку уровни транскрипции zag1 выше в пестиках, в то время как zmm2 экспрессируется в тычинках, что предполагает специализацию половых органов, которая объясняет отсутствие фенотипа в цветках-метелках zag1 (Mena и др. , 1996).
Паттерны экспрессии zag1 и zmm2 в значительной степени не перекрываются, поскольку уровни транскрипции zag1 выше в пестиках, в то время как zmm2 экспрессируется в тычинках, что предполагает специализацию половых органов, которая объясняет отсутствие фенотипа в цветках-метелках zag1 (Mena и др. , 1996).
Подобный результат был описан у риса. И OsMADS3 , ортолог zmm2 , и OSMADS58 , ортолог zag1 , экспрессируются в месте инициации тычинки и пестика.Мутации в OSMADS3 и OSMADS58 имеют последствия для спецификации органов, т.е. превращение тычинок в лодикулы и увеличение количества плодолистиков. Однако OSMADS58 , по-видимому, играет роль в детерминированности меристемы цветков, потому что мутанты постоянно имели неопределенное развитие органов. Кроме того, эффекты мутаций OSMADS3 на идентичность тычинок более серьезные, чем таковые мутаций OSMADS58 (Yamaguchi et al. , 2006).Хотя мутации в zmm2 еще не были выделены, в настоящее время имеются данные, указывающие на то, что, хотя zmm2 / OSMADS3 и zag1 / OSMADS58 вносят вклад в спецификацию тычинок и плодолистиков, их вклад неодинаков, причем первый более важен для идентичности тычинок, а последний — для правильного развития плодолистика и детерминированности цветочной меристемы.
, 2006).Хотя мутации в zmm2 еще не были выделены, в настоящее время имеются данные, указывающие на то, что, хотя zmm2 / OSMADS3 и zag1 / OSMADS58 вносят вклад в спецификацию тычинок и плодолистиков, их вклад неодинаков, причем первый более важен для идентичности тычинок, а последний — для правильного развития плодолистика и детерминированности цветочной меристемы.
Анализ двойных мутантов si1 и zag1 показывает ожидаемый фенотип для двойного мутанта BC, т.е.е. потеря лодикулы и идентичности тычинок и, вместо этого, образование нескольких мутовок органов, подобных цветковым/palea, что указывает на потерю определенности цветочной меристемы, а также на дефекты идентичности органов (Ambrose et al. , 2000).
Происхождение стерильных частей цветков трав остается загадкой на протяжении многих лет. В последние несколько лет стало очевидным, что модель развития цветков ABC применима с некоторыми модификациями к кукурузе и рису. Например, фенотипы мутаций в генах B- и C-классов кукурузы и их ортологи в рисе (Nagasawa et al., 2003), указывает на то, что лодикулы и лепестки имеют общего предка и развиваются в эквивалентных мутовках цветков травы и эвдикота соответственно (Irish, 2000). Интерпретация цветковых чешуй и палеа, однако, более сложна, потому что гомеотические преобразования у мутантов кукурузы B- и C-классов генерируют листовидные органы, которые имеют характеристики обоих (Ambrose et al. , 2000), хотя у риса мутации в SUPERWOMAN1 ( SPW1 ) превращают лодикулы в палеоподобные органы (Nagasawa et al., 2003). Одна из гипотез предполагает, что нижняя цветковая чешуя возникла в результате редукции и слияния прицветников, образовавшихся вне цветка у общего предка злаков и сестринских линий (Whipple and Schmidt, 2006).
Например, фенотипы мутаций в генах B- и C-классов кукурузы и их ортологи в рисе (Nagasawa et al., 2003), указывает на то, что лодикулы и лепестки имеют общего предка и развиваются в эквивалентных мутовках цветков травы и эвдикота соответственно (Irish, 2000). Интерпретация цветковых чешуй и палеа, однако, более сложна, потому что гомеотические преобразования у мутантов кукурузы B- и C-классов генерируют листовидные органы, которые имеют характеристики обоих (Ambrose et al. , 2000), хотя у риса мутации в SUPERWOMAN1 ( SPW1 ) превращают лодикулы в палеоподобные органы (Nagasawa et al., 2003). Одна из гипотез предполагает, что нижняя цветковая чешуя возникла в результате редукции и слияния прицветников, образовавшихся вне цветка у общего предка злаков и сестринских линий (Whipple and Schmidt, 2006).
Локусы количественных признаков, контролирующие развитие соцветий
Большая часть естественных вариаций формы соцветий, наблюдаемых у кукурузы и других видов трав, на самом деле обусловлена кумулятивным эффектом нескольких локусов. Поэтому, а также из-за экономической важности кукурузы и трав в целом изучение локусов количественных признаков (QTL) является важной областью генетики зерновых, направленной на повышение урожайности.В последнее время количественные исследования активизировались благодаря развитию геномных инструментов, таких как секвенирование генома риса и быстрое создание очень плотных генетических карт для нескольких видов трав. Как следствие, картирование QTL со значительно улучшенной разрешающей способностью теперь является мощным инструментом для выявления генов, контролирующих важные признаки. Недавно два сообщения охарактеризовали вклад QTL в архитектуру соцветий трав. Используя две инбредные линии сорго с разными соцветиями, Brown et al. (2006) картировали QTL по количеству ветвей от первого до третьего порядка, длине ветвей и диаметру позвоночника, среди прочих признаков. Их результаты предполагают, что ветви разных порядков находятся под контролем разных локусов. Два гена, Dwr3 ( br2 у кукурузы) и Sbra2 (ортолог кукурузы ra2 ), картированы в двух областях с QTL.
Поэтому, а также из-за экономической важности кукурузы и трав в целом изучение локусов количественных признаков (QTL) является важной областью генетики зерновых, направленной на повышение урожайности.В последнее время количественные исследования активизировались благодаря развитию геномных инструментов, таких как секвенирование генома риса и быстрое создание очень плотных генетических карт для нескольких видов трав. Как следствие, картирование QTL со значительно улучшенной разрешающей способностью теперь является мощным инструментом для выявления генов, контролирующих важные признаки. Недавно два сообщения охарактеризовали вклад QTL в архитектуру соцветий трав. Используя две инбредные линии сорго с разными соцветиями, Brown et al. (2006) картировали QTL по количеству ветвей от первого до третьего порядка, длине ветвей и диаметру позвоночника, среди прочих признаков. Их результаты предполагают, что ветви разных порядков находятся под контролем разных локусов. Два гена, Dwr3 ( br2 у кукурузы) и Sbra2 (ортолог кукурузы ra2 ), картированы в двух областях с QTL. Dw3 , который кодирует P-гликопротеин, ответственный за транспорт ауксина (Multani et al. , 2003), сопоставленный с QTL для высоты растения, длины стержня и ветви.Ген Sbra2 тесно совмещен с одним из двух QTL, обнаруженных по номеру первичной ветви.
Dw3 , который кодирует P-гликопротеин, ответственный за транспорт ауксина (Multani et al. , 2003), сопоставленный с QTL для высоты растения, длины стержня и ветви.Ген Sbra2 тесно совмещен с одним из двух QTL, обнаруженных по номеру первичной ветви.
Используя кукурузные метелки, Upadyayula и коллеги идентифицировали два QTL для большего количества ветвей, пять для плотности пар колосков на центральном колосе и два для плотности пар колосков на ветвях (Upadyayula et al. , 2006). Интересно, что последние два набора QTL (пары колосков на центральном шипе по сравнению с парами колосков на первичных ветвях) не перекрываются, что снова указывает на то, что разные локусы играют важную роль на разных стадиях развития.В початках были обнаружены QTL, которые контролируют количество зерен в ряду, плотность зерен, количество рядов в колосе, диаметр початка и массу початка. Некоторые гены, идентифицированные по мутантным фенотипам, совместно локализованы в QTL. К ним относятся ra1 , который точно соответствует QTL для номера ветви, td1 , который находится в той же области, что и QTL, которые контролируют массу початка, угол ветвления метелки и плотность пар колосков на первичных ветвях, ra2 , который картирует в регион с QTL для номера ядра на строку и fea2 , который локализуется в регион с QTL для номера ветви.Некоторые QTL были обнаружены в регионах с неизвестными генами, что указывает на то, что картирование QTL может помочь идентифицировать новые гены, участвующие в развитии соцветия (Upadyayula et al. , 2006).
К ним относятся ra1 , который точно соответствует QTL для номера ветви, td1 , который находится в той же области, что и QTL, которые контролируют массу початка, угол ветвления метелки и плотность пар колосков на первичных ветвях, ra2 , который картирует в регион с QTL для номера ядра на строку и fea2 , который локализуется в регион с QTL для номера ветви.Некоторые QTL были обнаружены в регионах с неизвестными генами, что указывает на то, что картирование QTL может помочь идентифицировать новые гены, участвующие в развитии соцветия (Upadyayula et al. , 2006).
Выводы
Травы являются самой важной сельскохозяйственной культурой в мире, но исследования, направленные на понимание механизмов органогенеза, были ограничены. Разработка новых методов в сочетании с мощью генетики кукурузы открыла новую эру в исследованиях биологии трав.Рис имеет полностью секвенированный геном, а кукуруза и сорго секвенируются. В настоящее время трансформация риса является обычным делом, и вскоре будет полностью секвенирован новый организм Brachypodium distachyon с очень коротким жизненным циклом и небольшим геномом (Draper et al. , 2001; Vogel et al. , 2005). ). Геномная синтения в семье успешно использовалась для клонирования генов кукурузы и пшеницы. Большинство генетических сетей детерминированности меристем и спецификации органов сохраняются у кукурузы и риса, за заметным исключением ra1 (Vollbrecht et al., 2005). Будущие эксперименты будут использовать синтению трав, чтобы разгадать механизмы, лежащие в основе изменений в архитектуре соцветий.
В настоящее время трансформация риса является обычным делом, и вскоре будет полностью секвенирован новый организм Brachypodium distachyon с очень коротким жизненным циклом и небольшим геномом (Draper et al. , 2001; Vogel et al. , 2005). ). Геномная синтения в семье успешно использовалась для клонирования генов кукурузы и пшеницы. Большинство генетических сетей детерминированности меристем и спецификации органов сохраняются у кукурузы и риса, за заметным исключением ra1 (Vollbrecht et al., 2005). Будущие эксперименты будут использовать синтению трав, чтобы разгадать механизмы, лежащие в основе изменений в архитектуре соцветий.
Рис. 2.
Кукуруза – однодомное растение с тычиночными цветками в кистях и пестичными цветками в колосьях. (A) Пара тычиночных цветков, на каждом из которых есть нижняя цветковая чешуя, палея и три тычинки (две лодикулы здесь не видны). (B) Ряд пестичных цветков, на каждом из которых виден один шелк (стиль). (C, D) Схематическое изображение, показывающее расположение органов цветения в паре тычиночных (C) и пестичных (D) колосков.
(C, D) Схематическое изображение, показывающее расположение органов цветения в паре тычиночных (C) и пестичных (D) колосков.
Рис. 2.
Кукуруза – однодомное растение с тычиночными цветками в кистях и пестичными цветками в колосьях. (A) Пара тычиночных цветков, на каждом из которых есть нижняя цветковая чешуя, палея и три тычинки (две лодикулы здесь не видны). (B) Ряд пестичных цветков, на каждом из которых виден один шелк (стиль). (C, D) Схематическое изображение, показывающее расположение органов цветения в паре тычиночных (C) и пестичных (D) колосков.
Рис. 3.
Соцветия кукурузы и мутанты цветения. (A-C) Мутанты ramosa имеют повышенную неопределенность боковых органов, трансформируя детерминированные меристемы (пары колосков) в ветви с различной степенью неопределенности. (А) ра1 . (Б) ра2 . (С) ра3 . (D) tasselseed4 ( ts4 ) представляет собой мутант, у которого не удается абортировать пестики в колосках-метелках, а также наблюдается усиленное ветвление. Обратите внимание на распространение шелка из-за образования пестика в кисточке (фото любезно предоставлено Джорджем Чаком). (Е) Ц6 также имеет феминизацию кисточек. На этом снимке шелка были удалены, чтобы показать формирование пестика. (F) Fascicled ear1 (Fas1) , мутант с фасциальным ухом и увеличенным числом рядов ядер (фото предоставлено China Lunde). (G) Мутант fuzzy tassel дает дополнительные цветки на колосок, несколько стерильных частей цветка и лишены нормальных чешуй.Показан сегмент мутантной кисточки слева по сравнению с диким типом справа.
Обратите внимание на распространение шелка из-за образования пестика в кисточке (фото любезно предоставлено Джорджем Чаком). (Е) Ц6 также имеет феминизацию кисточек. На этом снимке шелка были удалены, чтобы показать формирование пестика. (F) Fascicled ear1 (Fas1) , мутант с фасциальным ухом и увеличенным числом рядов ядер (фото предоставлено China Lunde). (G) Мутант fuzzy tassel дает дополнительные цветки на колосок, несколько стерильных частей цветка и лишены нормальных чешуй.Показан сегмент мутантной кисточки слева по сравнению с диким типом справа.
Рис. 3.
Мутанты соцветий и цветения кукурузы. (A-C) Мутанты ramosa имеют повышенную неопределенность боковых органов, трансформируя детерминированные меристемы (пары колосков) в ветви с различной степенью неопределенности. (А) ра1 . (Б) ра2 . (С) ра3 . (D) tasselseed4 ( ts4 ) представляет собой мутант, у которого не удается абортировать пестики в колосках-метелках, а также наблюдается усиленное ветвление. Обратите внимание на распространение шелка из-за образования пестика в кисточке (фото любезно предоставлено Джорджем Чаком). (Е) Ц6 также имеет феминизацию кисточек. На этом снимке шелка были удалены, чтобы показать формирование пестика. (F) Fascicled ear1 (Fas1) , мутант с фасциальным ухом и увеличенным числом рядов ядер (фото предоставлено China Lunde). (G) Мутант fuzzy tassel дает дополнительные цветки на колосок, несколько стерильных частей цветка и лишены нормальных чешуй.Показан сегмент мутантной кисточки слева по сравнению с диким типом справа.
Обратите внимание на распространение шелка из-за образования пестика в кисточке (фото любезно предоставлено Джорджем Чаком). (Е) Ц6 также имеет феминизацию кисточек. На этом снимке шелка были удалены, чтобы показать формирование пестика. (F) Fascicled ear1 (Fas1) , мутант с фасциальным ухом и увеличенным числом рядов ядер (фото предоставлено China Lunde). (G) Мутант fuzzy tassel дает дополнительные цветки на колосок, несколько стерильных частей цветка и лишены нормальных чешуй.Показан сегмент мутантной кисточки слева по сравнению с диким типом справа.
Работа была поддержана постдокторской стипендией NSF для EB и грантом NSF 0110189 для SH.
Каталожные номера
, , , , , .Молекулярный и генетический анализы гена silky1 выявили консервативность в спецификации органов цветка между эвдикотами и однодольными
,Molecular Cell
,2000
, vol.5
(стр.569
—579
), , , , , , .
Клонирование и характеристика кукурузы An1 Ген
,Растительная клетка
,1995
, vol.7
(стр.75
—84
), , , , , .Дубликат FLORICAULA/LEAFY гомологи zfl1 и zfl2 контролируют архитектуру соцветия и рисунок цветка кукурузы
,Развитие
,2003
, vol.130
(стр.2385
—2395
), , , , , , , .толстая кисточка dwarf1 кодирует предполагаемый кукурузный ортолог Arabidopsis CLAVATA1 богатая лейцином рецептороподобная киназа
,Development
,2005
, vol.132
(стр.1235
—1245
), , , .Генетика и эволюция соцветий трав и развитие цветков
,Физиология растений и клеток
,2005
, том.46
(стр.69
—78
), , , , , .ramosa2 кодирует белок домена латеральной границы органов, который определяет судьбу стволовых клеток в меристемах ветвей кукурузы
18
(стр.
574
—585
), , , , .Комплементарные цветочные гомеотипы являются результатом противоположной ориентации транспозона в локусе plena Antirrhinum
,Cell
,1993
, vol.72
(стр.85
—95
), , , , , .Наследование архитектуры соцветий у сорго
,Теоретическая и прикладная генетика
,2006
, том.113
(стр.931
—942
), .Гены гибели клеток и гены защиты клеток определяют судьбу пестиков у кукурузы
126
(стр.435
—441
), .Поддержание апикальной меристемы побегов: искусство динамического баланса
8
(стр.394
—401
), , , .Регуляция ауксинового ответа протеинкиназой PINOID
,Cell
,2000
, vol.100
(стр.469
—478
), , , и др.Ген FLORAL ORGAN NUMBER4 , кодирующий предполагаемого ортолога Arabidopsis CLAVATA3 , регулирует размер апикальной меристемы риса
142
(стр.
1039
—1052
), , .Контроль судьбы меристемы колоска кукурузы с помощью APETALA2 -подобного гена индетерминантного колоска1
,Гены и развитие
,1998
, vol.12
(стр.1145
—1154
), , , , .Контроль идентичности меристемы колосков разветвленным геном silkless1 у кукурузы
,Science
,2002
, vol.298
(стр.1238
—1241
), , .CLAVATA1 , регулятор меристемы и развития цветка Arabidopsis
,Development
,1993
, vol.119
(стр.397
—418
), , .CLAVATA3 – специфический регулятор развития побегов и цветочной меристемы, влияющий на те же процессы, что и CLAVATA1
,Развитие
,1995
, vol.121
(стр.2057
—2067
). , , , .Колосок и морфология цветков
,Систематика и эволюция трав
,1987
Вашингтон, округ Колумбия
Smithsonian Institution Press
(стр.
21
—070 9).
Война мутовок: генетические взаимодействия, контролирующие развитие цветка
,Природа
,1991
, том.353
(стр.31
—37
), , , , , .FLORICAULA : гомеозисный ген, необходимый для развития цветка у Antirrhinum majus
,Cell
,1990
, vol.63
(стр.1311
—1322
), , .Неопределенный ген кодирует белок цинкового пальца и регулирует генерируемый листьями сигнал, необходимый для перехода к цветению у кукурузы
,Cell
,1998
, vol.93
(стр.593
—603
), , , и др.Подробная карта взаимодействия транскрипционных факторов Arabidopsis MADS box
17
(стр.1424
—1433
), .Процесс определения пола у кукурузы
,Наука
,1994
, том.266
(стр.1501
—1505
), , .Ген определения пола TASSELSEED2 кукурузы кодирует короткоцепочечную алкогольдегидрогеназу, необходимую для аборта цветочных органов на определенной стадии
,Cell
,1993
, vol.
74
(стр.757
—768
), , , , , , .Brachypodium distachyon . Новая модельная система для функциональной геномики трав
,Физиология растений
,2001
, том.127
(стр.1539
—1555
), , , и др.PINOID-зависимый бинарный переключатель в апикально-базальном полярном нацеливании PIN направляет отток ауксина
306
(стр.862
—865
), , , , , , , .Роль бесплодного стебля1 в архитектуре кукурузы
,Природа
,2004
, том.432
(стр.630
—635
), , , , , , .Регуляция транспорта полярных ауксинов с помощью AtPIN1 в Arabidopsis сосудистой ткани
,Science
,1998
, vol.282
(стр.2226
—2230
).Филогения и подсемейная классификация злаков (Poaceae)
,Анналы Ботанического сада Миссури
,2001
, том.88
(стр.
373
—457
), .Генетика доминантной нечувствительной к гиббереллину карликовости кукурузы
,Генетика
,1989
, vol.121
(стр.827
—838
).Мутации семян метелок класса II свидетельствуют о множественных типах меристем соцветий у кукурузы (Poaceae)
84
(стр.1502
—1515
).Вариации на тему: развитие и эволюция цветка
,Биология генома
,2000
, том.1
(стр.1015.1
—1015.4
), , .Взаимодействия между генами метелки и другими определяющими пол генами кукурузы
,Генетика развития
,1994
, vol.15
(стр.155
—171
), .Развитие растений кукурузы из культивируемых верхушек побегов
,Planta
,1988
, vol.175
(стр.9
—12
), , .Ген Arabidopsis CLAVATA2 кодирует рецептороподобный белок, необходимый для стабильности рецептороподобной киназы CLAVATA1
,The Plant Cell
,1999
, vol.
11
(стр.1925
—1933
).Взаимодействие специфических генов, определяющих пол у раздельнополой кукурузы
,Труды Шестого Международного генетического конгресса
,1932
, том.2
(стр.104
—107
), .CLAVATA2 , регулятор развития меристемы и органов у Arabidopsis
,Development
,1998
, vol.125
(стр.3843
—3851
), , , .Мутации с потерей функции в гене гомеобокса кукурузы, узловатые1 , дефекты в поддержании меристемы побега
,Развитие
,1997
, vol.124
(стр.3045
—3054
), , , , , , , .LAX и SPA: основные регуляторы ветвления побегов у риса
100
(стр.11765
—11770
), , , , .ПУШИСТАЯ МЕТЕЛКА необходима для предотвращения образования пазушных меристем и установления идентичности цветочных меристем в колосках риса
,Развитие
,2003
, том.
130
(стр.3841
—3850
), .Развитие цветков и образование однополых колосков у Andropogoneae (Poaceae)
,American Journal of Botany
,1999
, vol.86
(стр.354
—366
), .бесплодное соцветие2 регулирует развитие пазушной меристемы соцветия кукурузы
,Развитие
,2001
, vol.128
(стр.2881
—2891
), , .Соцветие под любым другим названием: контроль идентичности меристемы кукурузы
5
(стр.61
—66
), , , , , .Диверсификация активности С-функции в развитии цветков кукурузы
,Наука
,1996
, том.274
(стр.1537
—1540
), , , , , .Потеря переносчика МЛУ в компактных стеблях кукурузы br2 и сорго dw 3 мутанты
,Science
,2003
, vol.302
(стр.81
—84
), , , , , , .
SUPERWOMAN1 и ПОНИЖАЮЩИЕ ЛИСТЬЯ гены контролируют идентичность органов цветка у риса
130
(стр.705
—718
), , . ,Мутанты кукурузы
,1997
Плейнвью, Нью-Йорк
Лаборатория Колд Спринг Харбор Пресс
Гены зеленой революции кодируют мутантные модуляторы ответа гиббереллина
,Nature
,1999
, vol.400
(стр.256
—261
).Реакция роста одногенных карликовых мутантов кукурузы на гибберелловую кислоту
42
(стр.185
—189
), .Изменения эндогенных гиббереллиноподобных веществ при реверсии пола верхушечных соцветий кукурузы
,Физиология растений
,1980
, том.66
(стр.793
—796
), , , , .Метаболический фермент трегалозы контролирует структуру соцветия кукурузы
,Nature
,2006
, vol.441
(стр.227
—230
), . ,
, Закономерности развития растений
,1989
Cambridge
Cambridge University Press
, , , , , . Ген НОМЕР ЦВЕТОЧНОГО ОРГАНА 1 регулирует размер цветочной меристемы риса и кодирует киназу богатого лейцином повторного рецептора, ортологичную Arabidopsis CLAVATA1 131 5649 5657 Ген фасциированного колоса2 кодирует богатый лейцином повторный рецептор-подобный белок, который регулирует пролиферацию меристемы побегов кукурузы 15 2755 2766 Генетический и QTL-анализ архитектуры метелок и початков кукурузы Theory of Applied Genetics 2006 112 592 606 Генетические подходы к развитию соцветий и листьев у кукурузы Молекулярные и клеточные основы формирования рисунка 1991 Кембридж, Англия Компания биологов, лтд. 105 112 Опосредованная Agrobacterium трансформация и развитие инбредных линий в модельной траве 84 199 211 Структура цветочных ветвей кукурузы и родственных трав Природа 2005 436 1119 1126 LEAFY контролирует идентичность цветочной меристемы в Arabidopsis Cell 1992 69 843 859 Сохранение функции цветочного гомеозисного гена класса B между кукурузой и Arabidopsis Развитие 2004 131 6083 6091 Генетика развития цветков травы Достижения в области ботанических исследований 2006 44 353 384 Функциональная диверсификация двух генов коробки MADS C-класса OSMADS3 и OSMADS58 в Oryza sativa The Plant Cell 2006 18 15 28 Белок, кодируемый гомеозисным геном Arabidopsis агамным , напоминает факторы транскрипции Nature 1990 346 35 39 © Автор [2007]. Опубликовано Oxford University Press [от имени Общества экспериментальной биологии]. Все права защищены. Для получения разрешений обращайтесь по электронной почте: [email protected]

Размер генома кукурузы (Zea mays L.), на который указывает обилие выступов длиной 180 п.н., связан со временем цветения) генетическое улучшение.
Растениеводство. 51 , 433–449 (2011).
Артикул Google Scholar
Prasanna, B.M. Разнообразие мировой гермоплазмы кукурузы: характеристика и использование. J. Biosci. 37 , 843–855 (2012).
КАС Статья пабмед Google Scholar
Кулешов Н. Н. Мировое разнообразие фенотипов кукурузы. Дж. Ам. соц. Агрон (1933 г.).
Realini, M. F., Poggio, L., Cámara-Hernandez, J. & Gonzalez, G. E. Внутривидовая изменчивость размера генома кукурузы: цитологические и фенотипические корреляции. АоБ Растения 8 , plv138 (2016).
Артикул Google Scholar
Чиа, Ж.-М. и др. .Maize HapMap2 идентифицирует существующие вариации изменяющегося генома. Нац. Жене. 44 , 803–807 (2012).
КАС Статья пабмед Google Scholar
Ян, X. и др. . Характеристика глобальной коллекции зародышевой плазмы и ее потенциальное использование для анализа сложных количественных признаков кукурузы. мол. Порода. 28 , 511–526 (2011).
мол. Порода. 28 , 511–526 (2011).
Артикул Google Scholar
Vigouroux, Y. и др. . Структура популяции и генетическое разнообразие рас кукурузы Нового Света, оцененные с помощью микросателлитов ДНК. утра. Дж. Бот. 95 , 1240–1253 (2008).
Артикул пабмед Google Scholar
Xia, X. C. и др. . Генетическое разнообразие инбредных линий кукурузы CIMMYT исследовано с помощью SSR-маркеров. Растениеводство. 44 , 2230–2237 (2004).
Артикул Google Scholar
Баклер Э. С., Гаут Б. С. и Макмаллен М. Д. Молекулярное и функциональное разнообразие кукурузы. Курс. мнение биол. растений 9 , 172–176 (2006).
КАС Статья пабмед Google Scholar
Springer, N.M. и др. . Инбредные сорта кукурузы демонстрируют высокие уровни вариации числа копий (CNV) и вариации присутствия/отсутствия (PAV) в содержании генома. Генетика PLoS. 5 , e1000734 (2009).
Артикул пабмед ПабМед Центральный Google Scholar
Muñoz-Diez, C., Vitte, C., Ross-Ibarra, J., Gaut, BS & Tenaillon, MI Использование следующего секвенирования для исследования вариаций размера генома и содержания мобильных элементов в Plant Transposable Elements (ред.Грандбастьен, М.-А. и Касакуберта, Дж. М.) 41–58 (Springer Berlin Heidelberg, 2012).
Beaulieu, J.M. и др. . Коррелированная эволюция размера генома и массы семян. Новый Фитол. 173 , 422–437 (2007).
Артикул пабмед Google Scholar
Sugiyama, S. Основа развития межвидовых различий в размере листьев и удельной площади листьев среди видов трав C3. Функц. Экол. 19 , 916–924 (2005).
Артикул Google Scholar
Tenaillon, M.I., Manicacci, D., Nicolas, S.D., Tardieu, F. & Welcker, C. Проверка связи между размером генома и скоростью роста кукурузы. PeerJ 4 , e2408 (2016).
Артикул пабмед ПабМед Центральный Google Scholar
Rayburn, A.L., Dudley, J.W. & Biradar, D.P. Отбор на раннее цветение приводит к одновременному отбору на пониженное содержание ядерной ДНК в кукурузе. Порода растений. 112 , 318–322 (1994).
Артикул Google Scholar

Díez, C.M. и др. . Изменение размера генома дикой и культивируемой кукурузы в зависимости от высотного градиента. Новый Фитол. 199 , 264–276 (2013).
Артикул пабмед ПабМед Центральный Google Scholar
Rayburn, A.L., Price, HJ, Smith, J.D. & Gold, J.R. Гетерохроматин C-диапазона и содержание ДНК в Zea mays . Являюсь. Дж. Бот. 72 , 1610–1617 (1985).
Артикул Google Scholar
Schnable, P. S. и др. .Геном кукурузы B73: сложность, разнообразие и динамика. Наука 326 , 1112–1115 (2009).
КАС Статья пабмед ОБЪЯВЛЕНИЯ Google Scholar
Пьегу, Б. и др. . Удвоение размера генома без полиплоидизации: динамика геномной экспансии, вызванной ретротранспозицией, у Oryza australiensis , дикого родственника риса. Рез. генома. 16 , 1262–1269 (2006).
Рез. генома. 16 , 1262–1269 (2006).
КАС Статья пабмед ПабМед Центральный Google Scholar
Ананьев, Е. В., Филлипс, Р. Л. и Райнс, Х. В. Тандемный повтор, связанный с выступом, у кукурузы, способный образовывать сегменты ДНК с загибом: являются ли выступы хромосом мегатранспозонами? Проц. Натл. акад. науч. США 95 , 10785–10790 (1998).
КАС Статья пабмед ПабМед Центральный ОБЪЯВЛЕНИЯ Google Scholar
McClintock, B. Морфология хромосом в Zea mays . Наука 69 , 629 (1929).
КАС Статья пабмед ОБЪЯВЛЕНИЯ Google Scholar
McClintock, B. Цитологическая демонстрация места взаимообмена между двумя негомологичными хромосомами Zea mays . Проц. Натл. акад. науч. США 16 , 791–796 (1930 г.).
Проц. Натл. акад. науч. США 16 , 791–796 (1930 г.).
КАС Статья пабмед ПабМед Центральный ОБЪЯВЛЕНИЯ Google Scholar
Tenaillon, M.I., Hufford, M.B., Gaut, B.S. & Ross-Ibarra, J. Размер генома и содержание мобильных элементов, определенные высокопроизводительным секвенированием кукурузы и Zea luxurians . Геном Биол. Эвол. 3 , 219–229 (2011).
КАС Статья пабмед ПабМед Центральный Google Scholar
Dennis, E.S. & Peacock, W.J. Гомология гетерохроматина Knob у кукурузы и ее родственников. Дж. Мол. Эвол. 20 , 341–350 (1984).
КАС Статья пабмед Google Scholar
Ананьев Е.В., Филипс Р.Л. и Райнс Х.W. Сложная структура ДНК выступа на хромосоме 9 кукурузы: инвазия ретротранспозона в гетерохроматин. Генетика 149 , 2025–2037 (1998).
Генетика 149 , 2025–2037 (1998).
КАС пабмед ПабМед Центральный Google Scholar
Kato, A., Lamb, J.C. & Birchler, J.A. Раскрашивание хромосом с использованием повторяющихся последовательностей ДНК в качестве зондов для идентификации соматических хромосом у кукурузы. Проц. Натл.акад. науч. США 101 , 13554–13559 (2004 г.).
КАС Статья пабмед ПабМед Центральный ОБЪЯВЛЕНИЯ Google Scholar
Коласанти, Дж. и Мушински, М. Цветочный переход кукурузы в Справочнике кукурузы: ее биология (под редакцией Беннетцен, Дж. Л. и Хейк, С. К.) 41–55 (Springer New York, 2009).
Сальви, С. и др. . Консервативные некодирующие геномные последовательности, связанные с локусом количественного признака времени цветения у кукурузы. Проц. Натл. акад. науч. США 104 , 11376–11381 (2007 г.).
Проц. Натл. акад. науч. США 104 , 11376–11381 (2007 г.).
КАС Статья пабмед ПабМед Центральный ОБЪЯВЛЕНИЯ Google Scholar
Менг, X., Мушинский, М. Г. и Данилевская, О. Н. Ген FT -Like ZCN8 действует как цветочный активатор и участвует в чувствительности к фотопериоду у кукурузы. Растительная клетка 23 , 942–960 (2011).
КАС Статья пабмед ПабМед Центральный Google Scholar
Kozaki, A., Hake, S. & Colasanti, J. Регулятор времени цветения кукурузы ID1 представляет собой белок цинковых пальцев с новыми свойствами связывания ДНК. Рез. нуклеиновых кислот. 32 , 1710–1720 (2004).
КАС Статья пабмед ПабМед Центральный Google Scholar

Хунг, Х.-Ю. и др. . ZmCCT и генетические основы адаптации к длине дня, лежащие в основе распространения кукурузы после одомашнивания. Проц. Натл. акад. науч. США 109 , 1913–1921 (2012).
Артикул Google Scholar
Ян, Q. и др. . CACTA-подобный мобильный элемент в ZmCCT ослаблял чувствительность к фотопериоду и ускорял распространение кукурузы после одомашнивания. Проц. Натл. акад. науч. США 110 , 16969–16974 (2013 г.).
КАС Статья пабмед ПабМед Центральный ОБЪЯВЛЕНИЯ Google Scholar
Сюй, Дж. и др. . Генетическая архитектура времени цветения и чувствительности к фотопериоду у кукурузы, выявленная в обзоре QTL и метаанализе. Дж. Интегр. биол. растений 54 , 358–373 (2012).
КАС Статья пабмед Google Scholar
Buckler, ES и др. . Генетическая архитектура времени цветения кукурузы. Наука 325 , 714–718 (2009).
КАС Статья пабмед ОБЪЯВЛЕНИЯ Google Scholar
Ли, Х. и др. . Полногеномное ассоциативное исследование анализирует генетическую архитектуру биосинтеза масла в зернах кукурузы. Нац. Жене. 45 , 43–50 (2013).
КАС Статья пабмед Google Scholar
Тянь Ф. и др. . Полногеномное ассоциативное исследование архитектуры листа в популяции картирования вложенных ассоциаций кукурузы. Нац. Жене. 43 , 159–162 (2011).
КАС Статья пабмед Google Scholar

Лю, С. и др. .Полногеномный анализ генов ZmDREB и их связь с естественной изменчивостью засухоустойчивости на стадии проростков Zea mays L. PLoS Genet. 9 , e1003790 (2013 г.).
КАС Статья пабмед ПабМед Центральный Google Scholar
Мао, Х. и др. . Мобильный элемент в гене NAC связан с засухоустойчивостью проростков кукурузы. Нац. коммун. 6 , 8326 (2015).
КАС Статья пабмед ПабМед Центральный Google Scholar
Ян Дж. и др. . Редкая генетическая вариация Zea mays crtRB1 повышает уровень β-каротина в зерне кукурузы. Нац. Жене. 42 , 322–327 (2010).
КАС Статья пабмед Google Scholar

Сюй, К. и др. . Разработка массива 55K SNP кукурузы с улучшенным охватом генома для молекулярной селекции. мол. Порода. 37 , 20 (2017).
Артикул пабмед ПабМед Центральный Google Scholar
Ли, Ю. и др. . Идентификация генетических вариантов, связанных со временем цветения кукурузы, с использованием чрезвычайно большой мультигенетической фоновой популяции. Завод Ж. 86 , 391–402 (2016).
КАС Статья пабмед Google Scholar
de Aguiar-Perecin, M.L. & Vosa, C.G. С-полосатость кукурузы II. Идентификация соматических хромосом. Наследственность 54 , 37–42 (1985).
Артикул Google Scholar
Zhang, H. & Dawe, R. K. Общий размер центромер и размер генома тесно связаны между собой у десяти видов трав. Хромосомный Рез. 20 , 403–412 (2012).
Хромосомный Рез. 20 , 403–412 (2012).
КАС Статья пабмед ПабМед Центральный Google Scholar
Инициатива по изучению генома арабидопсиса. Анализ последовательности генома цветкового растения Arabidopsis thaliana . Природа 408 , 796–815 (2000).
Артикул ОБЪЯВЛЕНИЯ Google Scholar
Джин В. и др. . Молекулярная и функциональная диссекция центромеры В-хромосомы кукурузы. Растительная клетка 17 , 1412–1423 (2005).
КАС Статья пабмед ПабМед Центральный Google Scholar
Энке, Н. и др. . Сдвиги размера генома: эволюция кариотипа в секции Crepis Neglectoides (Asteraceae). Растение Биол. 17 , 775–786 (2015).
17 , 775–786 (2015).
КАС Статья пабмед Google Scholar
Doležel, J., Greilhuber, J. & Suda, J. Оценка содержания ядерной ДНК в растениях с помощью проточной цитометрии. Нац. протокол 2 , 2233–2244 (2007).
Артикул пабмед Google Scholar
Мюррей, М. Г. и Томпсон, В. Ф. Быстрое выделение ДНК растений с высокой молекулярной массой. Рез. нуклеиновых кислот. 8 , 4321–4326 (1980).
КАС Статья пабмед ПабМед Центральный Google Scholar
Перепел, Массачусетс и др. . Рассказ о трех платформах для секвенирования следующего поколения: сравнение секвенаторов Ion Torrent, Pacific Biosciences и Illumina MiSeq. БМС Геномика 13 , 341 (2012).
КАС Статья пабмед ПабМед Центральный Google Scholar
Bolger, A.M., Lohse, M. & Usadel, B. Trimmomatic: гибкий триммер для данных последовательностей Illumina. Биоинформатика 30 , 2114–2120 (2014).
КАС Статья пабмед ПабМед Центральный Google Scholar
Mortazavi, A., Williams, B.A., McCue, K., Schaeffer, L. & Wold, B. Картирование и количественная оценка транскриптомов млекопитающих с помощью RNA-Seq. Нац. Методы 5 , 621–628 (2008).
КАС Статья пабмед Google Scholar
Брэдбери, П. Дж. и др. . TASSEL: программное обеспечение для картирования ассоциаций сложных признаков в различных образцах. Биоинформатика 23 , 2633–2635 (2007).
КАС Статья пабмед Google Scholar
Ван, С., Дворкин, Д. и Да, Ю. SNPEVG: графический инструмент для построения графиков GWAS с помощью щелчков мыши. Биоинформатика BMC 13 , 319 (2012).
Артикул Google Scholar
ZmCCT регулирует зависящее от фотопериода цветение и реакцию кукурузы на стресс | BMC Plant Biology
Matsuoka Y, Vigouroux Y, Goodman MM, Sanchez GJ, Buckler E, Doebley J: Одноместное одомашнивание кукурузы, показанное мультилокусным микросателлитным генотипированием. Proc Natl Acad Sci 2002, 99(9):6080–6084.
Abou-Elwafa SF, Büttner B, Chia T, Schulze-Buxloh G, Hohmann U, Mutasa-Göttgens E, et al. Консервация и дивергенция генов автономного пути в регуляторной сети цветения Beta vulgaris. J Опытный бот. 2011;62(10):3359–74.
КАС пабмед Статья Google Scholar

Бюттнер Б., Абу-Эльвафа С.Ф., Чжан В., Юнг С., Мюллер А.Е. Обзор двухлетних мутантов Beta vulgaris, индуцированных ЭМС, выявил новый локус стрелковости, который не связан с геном стрелковости B. Theor Appl Genet. 2010;121(6):1117–31.
ПабМед Статья Google Scholar
Jin M, Liu X, Jia W, Liu H, Li W, Peng Y, et al. ZmCOL3, ген CCT, подавляет цветение кукурузы, вмешиваясь в циркадные часы и активируя экспрессию ZmCCT.J Integr Plant Biol. 2018;60(6):465–80.
КАС пабмед Статья Google Scholar
Hung H-Y, Shannon LM, Tian F, Bradbury PJ, Chen C, Flint-Garcia SA, et al. ZmCCT и генетическая основа адаптации к длине дня, лежащая в основе распространения кукурузы после одомашнивания. Proc Natl Acad Sci. 2012;109(28):E1913–21.
КАС пабмед ПабМед Центральный Статья Google Scholar

Yang Q, Li Z, Li W, Ku L, Wang C, Ye J и др. CACTA-подобный мобильный элемент в ZmCCT ослаблял чувствительность к фотопериоду и ускорял распространение кукурузы после одомашнивания. Proc Natl Acad Sci. 2013;110(42):16969–74.
КАС пабмед ПабМед Центральный Статья Google Scholar
Huang C, Sun H, Xu D, Chen Q, Liang Y, Wang X, et al. ZmCCT9 усиливает адаптацию кукурузы к более высоким широтам. Proc Natl Acad Sci U S A.2018;115(2):E334–e341.
КАС пабмед Статья Google Scholar
Guo L, Wang X, Zhao M, Huang C, Li C, Li D, et al. Ступенчатые цис-регуляторные изменения в ZCN8 способствуют адаптации кукурузы к времени цветения. Текущая биология. 2018;28(18):3005–15 e3004.
КАС пабмед Статья Google Scholar
Мэн Х, Мушинский М.Г., Данилевская О.Н.FT-подобный ген ZCN8 действует как цветочный активатор и участвует в чувствительности к фотопериоду у кукурузы. Растительная клетка. 2011;23(3):942–60.
Растительная клетка. 2011;23(3):942–60.
КАС пабмед ПабМед Центральный Статья Google Scholar
Liang Y, Liu Q, Wang X, Huang C, Xu G, Hey S, et al. ZmMADS69 действует как активатор цветения через регуляторный модуль ZmRap2.7-ZCN8 и способствует адаптации времени цветения кукурузы. Новый Фитол. 2019;221(4):2335–47.
КАС пабмед Статья Google Scholar
Данилевская О.Н., Мэн Х., Селинжер Д.А., Дешам С., Хермон П., Вансант Г. и соавт. Участие гена MADS-box ZMM4 в индукции цветения и развитии соцветий кукурузы. Завод Физиол. 2008;147(4):2054–69.
КАС пабмед ПабМед Центральный Статья Google Scholar
Яновский М.Ю., Кей С.А.Жизнь по календарю: как растения узнают, когда цвести. Nat Rev Mol Cell Biol. 2003;4(4):265–76.
КАС пабмед Статья Google Scholar

Corbesier L, Coupland G. Фотопериодическое цветение арабидопсиса: интеграция генетических и физиологических подходов к характеристике цветочного стимула. Окружающая среда растительной клетки. 2005;28(1):54–66.
КАС Статья Google Scholar
Wang X, Wu L, Zhang S, Wu L, Ku L, Wei X и другие. Надежная экспрессия и связь ZmCCA1 с циркадными ритмами у кукурузы. Отчет о растительных клетках 2011;30(7):1261–72.
ПабМед Статья КАС Google Scholar
Shi Y, Zhao X, Guo S, Dong S, Wen Y, Han Z, et al. ZmCCA1a на хромосоме 10 кукурузы задерживает цветение Arabidopsis thaliana. Фронт завод науч. 2020;11:78.
ПабМед ПабМед Центральный Статья Google Scholar
Kumimoto RW, Zhang Y, Siefers N, Holt BF 3rd. NF-YC3, NF-YC4 и NF-YC9 необходимы для CONSTANS-опосредованного цветения, зависящего от фотопериода, у Arabidopsis thaliana. Плант Дж. 2010;63(3):379–91.
Плант Дж. 2010;63(3):379–91.
КАС пабмед Статья Google Scholar
Цао С., Кумимото Р.В., Гнесутта Н., Калоджеро А.М., Мантовани Р., Холт Б.Ф. 3-й. Дистальный комплекс CCAAT/NUCLEAR FACTOR Y способствует формированию петли хроматина на промоторе FLOWERING LOCUS T и регулирует время цветения у арабидопсиса.Растительная клетка. 2014;26(3):1009–17.
КАС пабмед ПабМед Центральный Статья Google Scholar
Хван Ю.Х., Ким С.К., Ли К.С., Чанг Ю.С., Ли Д.Х., Ким Д.К. Функциональная консервация белков OsNF-YB/YC риса и AtNF-YB/YC арабидопсиса в регуляции времени цветения. Отчет о растительных клетках 2016;35(4):857–65.
КАС пабмед Статья Google Scholar
Su H, Cao Y, Ku L, Yao W, Cao Y, Ren Z и др. Двойные функции ZmNF-YA3 в реакциях цветения, зависящих от фотопериода, и реакции на абиотический стресс у кукурузы. J Опытный бот. 2018;69(21):5177–89.
J Опытный бот. 2018;69(21):5177–89.
КАС пабмед Статья Google Scholar
Тянь Л., Чжао С., Лю Х., Ку Л., Ван С., Хань З. и др. Альтернативный сплайсинг ZmCCA1 опосредует реакцию тропической кукурузы на засуху. ПЛОС Один. 2019;14(1):e0211623.
КАС пабмед ПабМед Центральный Статья Google Scholar
Лю Т., Карлссон Дж., Такеучи Т., Ньютон Л., Фарре Э.М. Прямая регуляция абиотических реакций с помощью компонента циркадных часов Arabidopsis PRR7. Плант Дж. 2013;76(1):101–14.
КАС пабмед Google Scholar
Wang W, Barnaby JY, Tada Y, Li H, Tör M, Caldelari D. Lee D-u, Fu X-D, Dong X: синхронизация иммунных реакций растений центральным циркадным регулятором. Природа. 2011;470(7332):110–4.
КАС пабмед ПабМед Центральный Статья Google Scholar

Мин Дж. Х., Чанг Дж. С., Ли К. Х., Ким К. С. CONSTANS-подобный фактор транскрипции 4, AtCOL4, положительно регулирует толерантность к абиотическому стрессу у арабидопсиса зависимым от абсцизовой кислоты образом. J Integr Plant Biol. 2015;57(3):313–24.
КАС пабмед Статья Google Scholar
Xue W, Xing Y, Weng X, Zhao Y, Tang W, Wang L, et al. Естественная изменчивость Ghd7 является важным регулятором срока колошения и потенциальной урожайности риса.Нат Жене. 2008;40(6):761–7.
КАС Статья Google Scholar
Ku L, Tian L, Su H, Wang C, Wang X, Wu L, et al. Двойные функции ZmCCT-ассоциированного локуса количественных признаков в реакциях цветения и стресса в условиях длинного дня. BMC Растение Биол. 2016;16(1):239.
ПабМед ПабМед Центральный Статья КАС Google Scholar

Chettoor AM, Givan SA, Cole RA, Coker CT, Unger-Wallace E, Vejlupkova Z, et al.Открытие новых транскриптов и гаметофитных функций с помощью анализа секвенирования РНК гаметофитных транскриптомов кукурузы. Геном биол. 2014;15(7):414.
ПабМед ПабМед Центральный Статья КАС Google Scholar
Wang B, Tseng E, Regulski M, Clark TA, Hon T, Jiao Y, et al. Выявление сложности транскриптома кукурузы с помощью долговременного секвенирования одной молекулы. Нац коммун. 2016;7(1):11708.
КАС пабмед ПабМед Центральный Статья Google Scholar
Tian T, Liu Y, Yan H, You Q, Yi X, Du Z и другие. agriGO v2.0: набор инструментов для анализа GO для сельскохозяйственного сообщества, обновление 2017 г. Нуклеиновые Кислоты Res. 2017;45(Н1):В122–н129.
КАС пабмед ПабМед Центральный Статья Google Scholar

Gendron JM, Pruneda-Paz JL, Doherty CJ, Gross AM, Kang SE, Kay SA. Белок циркадных часов арабидопсиса, TOC1, представляет собой ДНК-связывающий фактор транскрипции. Proc Natl Acad Sci. 2012;109(8):3167–72.
КАС пабмед ПабМед Центральный Статья Google Scholar
Song K, Kim HC, Shin S, Kim KH, Moon JC, Kim JY и др. Транскриптомный анализ генов времени цветения в условиях стресса засухи в листьях кукурузы. Фронт завод науч. 2017;8:267.
ПабМед ПабМед Центральный Google Scholar
Wang J, Ding H, Zhang A, Ma F, Cao J, Jiang M. Новый ген митоген-активируемой протеинкиназы кукурузы (Zea mays), ZmMPK3, участвует в реакции на различные сигналы окружающей среды.J Integr Plant Biol. 2010;52(5):442–52.
КАС пабмед Google Scholar
Wang B, Liu C, Zhang D, He C, Zhang J, Li Z. Влияние стрессовой реакции органов кукурузы на засуху на урожайность по данным транскриптомного анализа. BMC Растение Биол. 2019;19(1):335.
Влияние стрессовой реакции органов кукурузы на засуху на урожайность по данным транскриптомного анализа. BMC Растение Биол. 2019;19(1):335.
ПабМед ПабМед Центральный Статья КАС Google Scholar
Накаи Ю., Накахира Ю., Сумида Х., Такебаяси К., Нагасава Ю., Ямасаки К. и др.Факторы транскрипции белка одного цинкового пальца 1/2 сосудистых растений регулируют реакцию абиотического и биотического стресса у арабидопсиса. Плант Дж. 2013;73(5):761–75.
КАС пабмед Статья Google Scholar
Cordenonsi M, Zanconato F, Azzolin L, Forcato M, Rosato A, Frasson C, et al. Преобразователь гиппопотама TAZ придает клеткам рака молочной железы черты, связанные со стволовыми клетками рака. Клетка. 2011;147(4):759–72.
КАС пабмед ПабМед Центральный Статья Google Scholar

Overholtzer M, Zhang J, Smolen GA, Muir B, Li W, Sgroi DC, et al. Трансформирующие свойства YAP, кандидата в онкогены на ампликоне хромосомы 11q22. Proc Natl Acad Sci U S A. 2006;103(33):12405–10.
КАС пабмед ПабМед Центральный Статья Google Scholar
Ким М., Ким Т., Джонсон Р.Л., Лим Д.С. Транскрипционная корепрессорная функция преобразователей пути гиппопотама YAP и TAZ. Cell Rep. 2015;11(2):270–82.
КАС пабмед Статья Google Scholar
Lai X, Yan L, Lu Y, Schnable JC. Наборы генов, в значительной степени несцепленные, нацелены на отбор фенотипов синдрома одомашнивания у кукурузы и сорго. Плант Дж. 2018;93(5):843–55.
КАС пабмед Статья Google Scholar
Du X, Wang X, Zhang J, Zhen S, Liu Y, Fu J и др. Транскриптомный анализ лиственных почти изогенных линий дает молекулярное представление о цветочном переходе у кукурузы (Zea mays). Порода растений. 2020;139(5):883–91.
Порода растений. 2020;139(5):883–91.
КАС Статья Google Scholar
Lai X, Bendix C, Yan L, Zhang Y, Schnable JC, Harmon FG. Межвидовой анализ регуляции суточного гена у паникоидов выявляет известные и новые регуляторные мотивы. Геномика BMC. 2020;21(1):428.
КАС пабмед ПабМед Центральный Статья Google Scholar
Rawat R, Takahashi N, Hsu PY, Jones MA, Schwartz J, Salemi MR, et al.REVEILLE8 и PSEUDO-REPONSE REGULATOR5 образуют петлю отрицательной обратной связи в циркадных часах арабидопсиса. Генетика PLoS. 2011;7(3):e1001350.
КАС пабмед ПабМед Центральный Статья Google Scholar
Hsu PY, Devisetty UK, Harmer SL. Точность хронометража контролируется активатором цикла в арабидопсисе. электронная жизнь. 2013;2:e00473.
ПабМед ПабМед Центральный Статья КАС Google Scholar

Песня Н., Сюй З., Ван Дж., Цинь Ц., Цзян Х., Си В. и др. Полногеномный анализ семейства генов CONSTANS-LIKE кукурузы и профилирование экспрессии при обработке светом/темнотой и абсцизовой кислотой. Ген. 2018; 673:1–11.
КАС пабмед Статья Google Scholar
Minow MAA, Ávila LM, Turner K, Ponzoni E, Mascheretti I, Dussault FM, et al. Различные генные сети модулируют цветочную индукцию автономной кукурузы и теосинта, зависящего от фотопериода.J Опытный бот. 2018;69(12):2937–52.
КАС пабмед ПабМед Центральный Статья Google Scholar
Cheng XF, Wang ZY. Сверхэкспрессия COL9, гена CONSTANS-LIKE, задерживает цветение за счет снижения экспрессии CO и FT у Arabidopsis thaliana. Плант Дж. 2005; 43 (5): 758–68.
КАС пабмед Статья Google Scholar
Wu W, Zheng XM, Lu G, Zhong Z, Gao H, Chen L, et al. Связь функциональных полиморфизмов нуклеотидов в DTh3 с распространением выращивания риса на север в Азии. Proc Natl Acad Sci U S A. 2013;110(8):2775–80.
Связь функциональных полиморфизмов нуклеотидов в DTh3 с распространением выращивания риса на север в Азии. Proc Natl Acad Sci U S A. 2013;110(8):2775–80.
КАС пабмед ПабМед Центральный Статья Google Scholar
Bendix C, Marshall CM, Harmon FG. Гены циркадных часов повсеместно контролируют ключевые сельскохозяйственные признаки. Мол завод. 2015;8(8):1135–52.
КАС пабмед Статья Google Scholar
Farré EM, Liu T. Семейство регуляторов транскрипции PRR отражает сложность и эволюцию циркадных часов растений. Curr Opin Plant Biol. 2013;16(5):621–9.
ПабМед Статья КАС Google Scholar
Hajdu A, Dobos O, Domijan M, Balint B, Nagy I, Nagy F, et al. УДЛИНЕННЫЙ ГИПОКОТИЛ 5 обеспечивает передачу сигналов синим светом циркадным часам арабидопсиса. Плант Дж. 2018;96(6):1242–54.
КАС пабмед Статья Google Scholar
Саломе П.А., То Дж.П., Кибер Дж.Дж., МакКланг К.Р. Регуляторы ответа арабидопсиса ARR3 и ARR4 играют независимую от цитокининов роль в контроле циркадианного периода. Растительная клетка. 2006;18(1):55–69.
ПабМед ПабМед Центральный Статья КАС Google Scholar
Martí Ruiz MC, Hubbard KE, Gardner MJ, Jung HJ, Aubry S, Hotta CT, et al. Суточные колебания цитозольного свободного кальция регулируют циркадные часы арабидопсиса.Природа растений. 2018;4(9):690–8.
ПабМед Статья КАС Google Scholar
Kevei E, Gyula P, Fehér B, Tóth R, Viczián A, Kircher S, et al. Циркадные часы Arabidopsis thaliana регулируются малой ГТФазой LIP1. Карр Биол. 2007;17(17):1456–64.
КАС пабмед Статья Google Scholar

Li A, Li G, Zhao Y, Meng Z, Zhao M, Li C, et al.Комбинированный анализ экспрессии малых РНК и генов выявил роль микроРНК в реакции кукурузы на инфекцию вируса черной карликовости риса. Научный доклад 2018; 8 (1): 13502.
ПабМед ПабМед Центральный Статья КАС Google Scholar
Ши Ф., Чжан Ю., Ван К., Мэн К., Лю С., Ма Л. и др. Анализ профиля экспрессии кукурузы в ответ на Setosphaeria turcica. Ген. 2018;659:100–8.
КАС пабмед Статья Google Scholar
Yu Y, Shi J, Li X, Liu J, Geng Q, Shi H и др. Анализ транскриптома раскрывает молекулярные механизмы защитного ответа кукурузы на серую пятнистость листьев. Геномика BMC. 2018;19(1):742.
КАС пабмед ПабМед Центральный Статья Google Scholar
Walley JW, Shen Z, McReynolds MR, Schmelz EA, Briggs SP. Вызванное грибками гиперацетилирование белка в кукурузе, идентифицированное с помощью анализа ацетилома. Proc Natl Acad Sci U S A.2018;115(1):210–5.
Вызванное грибками гиперацетилирование белка в кукурузе, идентифицированное с помощью анализа ацетилома. Proc Natl Acad Sci U S A.2018;115(1):210–5.
КАС пабмед Статья Google Scholar
Llorente F, Alonso-Blanco C, Sánchez-Rodriguez C, Jorda L, Molina A. Киназа, подобная рецептору ERECTA, и гетеротримерный G-белок арабидопсиса необходимы для устойчивости к некротрофному грибку Plectosphaerella cucumerina. Плант Дж. 2005; 43 (2): 165–80.
КАС пабмед Статья Google Scholar
Ю Г.Л., Катагири Ф., Осубел Ф.М. Мутации арабидопсиса в локусе RPS2 приводят к потере устойчивости к штаммам pseudomonas syringae, экспрессирующим ген авирулентности avrRpt2. Mol Взаимодействие растений и микробов. 1993;6(4):434–43.
КАС Статья Google Scholar
Кункель Б.Н., Бент А.Ф., Дальбек Д. , Иннес Р.В., Стаскавич Б.Дж. RPS2, локус устойчивости к болезням арабидопсиса, определяющий распознавание штаммов pseudomonas syringae, экспрессирующих ген авирулентности avrRpt2.Растительная клетка. 1993;5(8):865–75.
, Иннес Р.В., Стаскавич Б.Дж. RPS2, локус устойчивости к болезням арабидопсиса, определяющий распознавание штаммов pseudomonas syringae, экспрессирующих ген авирулентности avrRpt2.Растительная клетка. 1993;5(8):865–75.
КАС пабмед ПабМед Центральный Google Scholar
Рохас К.М., Сентил-Кумар М., Ван К., Рю К.М., Каундал А., Майсур К.С. Гликолатоксидаза модулирует передачу сигнала, опосредованную активными формами кислорода, во время устойчивости, не являющейся хозяином, у Nicotiana benthamiana и Arabidopsis. Растительная клетка. 2012;24(1):336–52.
КАС пабмед ПабМед Центральный Статья Google Scholar
Wu Q, Xu F, Liu L, Char SN, Ding Y, Je BI и др. Субъединица β гетеротримерного G-белка кукурузы контролирует развитие меристемы побегов и иммунные реакции. Proc Natl Acad Sci U S A. 2020;117(3):1799–805.
КАС пабмед Статья Google Scholar

Li H, Huang K, Du H, Wang H, Chen X, Gao S, et al. Полногеномный анализ корепрессоров семейства Gro/Tup1 и их реакции на гормоны и абиотические стрессы у кукурузы.Дж. Биол. растения. 2016;59(6):603–15.
КАС Статья Google Scholar
Colombo L, Marziani G, Masiero S, Wittich PE, Schmidt RJ, Gorla MS, et al. BRANCHED SILKLESS опосредует переход от колоска к цветочной меристеме во время развития колоса Zea mays. Плант Дж. 1998;16(3):355–63.
КАС Статья Google Scholar
Xiao W, Custard KD, Brown RC, Lemmon BE, Harada JJ, Goldberg RB, et al.Метилирование ДНК имеет решающее значение для эмбриогенеза арабидопсиса и жизнеспособности семян. Растительная клетка. 2006;18(4):805–14.
КАС пабмед ПабМед Центральный Статья Google Scholar
Yamauchi T, Johzuka-Hisatomi Y, Terada R, Nakamura I, Iida S. Ген MET1b, кодирующий поддерживающую ДНК-метилтрансферазу, необходим для нормального развития риса. Завод Мол Биол. 2014;85(3):219–32.
Ген MET1b, кодирующий поддерживающую ДНК-метилтрансферазу, необходим для нормального развития риса. Завод Мол Биол. 2014;85(3):219–32.
КАС пабмед Статья Google Scholar
Сюй Дж., Чжан Х.И., Се Ч., Сюэ Х.В., Дийхуис П., Лю К.М. ЭМБРИОНАЛЬНЫЙ ФАКТОР 1 кодирует AMP-дезаминазу и необходим для перехода зиготы в эмбрион у арабидопсиса. Плант Дж. 2005;42(5):743–56.
КАС пабмед Статья Google Scholar
Селесник Х., Али Г.С., Робисон Ф.М., Редди А.С. Факторы транскрипции Arabidopsis thaliana VOZ (сосудистые растения с одним цинковым пальцем) необходимы для правильной регуляции времени цветения.Биология открытая. 2013;2(4):424–31.
КАС пабмед ПабМед Центральный Статья Google Scholar
Cao Y, Zeng H, Ku L, Ren Z, Han Y, Su H, et al. ZmIBh2-1 регулирует архитектуру растений кукурузы. J Опытный бот. 2020;71(10):2943–55.
КАС пабмед ПабМед Центральный Статья Google Scholar
Тивари С.Б., Ван XJ, Хаген Г., Гилфойл Т.Дж.Белки AUX/IAA являются активными репрессорами, и их стабильность и активность модулируются ауксином. Растительная клетка. 2001;13(12):2809–22.
КАС пабмед ПабМед Центральный Статья Google Scholar
O’Malley RC, Huang SC, Song L, Lewsey MG, Bartlett A, Nery JR, et al. Особенности цистрома и эпистрома формируют регуляторный ландшафт ДНК. Клетка. 2016;165(5):1280–92.
КАС пабмед ПабМед Центральный Статья Google Scholar
Лангмид Б., Зальцберг С.Л. Быстрое выравнивание с промежутками чтения с помощью галстука-бабочки 2. Nat Methods. 2012;9(4):357–9.
КАС пабмед ПабМед Центральный Статья Google Scholar
Zhang Y, Liu T, Meyer CA, Eeckhoute J, Johnson DS, Bernstein BE, et al. Модельный анализ ChIP-Seq (MACS). Геном биол. 2008;9(9):R137.
ПабМед ПабМед Центральный Статья КАС Google Scholar
Вималанатан К., Фридберг И., Андорф С.М., Лоуренс-Дилл С.Дж. Методы аннотирования, оценка и обзор Maize GO (maize-GAMER). Завод прямой. 2018;2(4):e00052.
ПабМед ПабМед Центральный Статья КАС Google Scholar
Benjamini Y, Yekutieli D. Контроль частоты ложных открытий при множественном тестировании в условиях зависимости. Энн Стат. 2001;29(4):1165 -1188, 1124.
Статья Google Scholar
Zhang H, Xiang Y, He N, Liu X, Liu H, Fang L и др. Повышенная выработка витамина С, опосредованная АБК-индуцированной РТР-подобной нуклеотидазой, улучшает засухоустойчивость арабидопсиса и кукурузы. Мол завод. 2020;13(5):760–76.
КАС пабмед Статья Google Scholar
Квасьневски М., Дашковска-Голец А., Яняк А., Хвилковска К., Новаковска Ю., Саблок Г., и другие. Анализ транскриптома показывает роль корневых волосков как датчиков окружающей среды для поддержания функций растений в условиях дефицита воды.J Опытный бот. 2016;67(4):1079–94.
КАС пабмед Статья Google Scholar
Спустенное СНГ-регуляторные изменения в ZCN8. Способствуют в ZCN8. Способствуют в кукурузе. Лежит в основе крупных QTL для цветения Время цветения, обнаруженные в популяции кукурузы-теозинта
Два независимых CIS -Variants в ZCN8 Промоутер Acception Action ZCN8 Время цветения Кукурузы
Две СНГ — 2 в ZCN8 возник поэтапно, чтобы усилить адаптацию кукурузы
Доколумбовое распространение кукурузы включало многочисленные локусы с отчетливыми особенностями mays ) был одомашнен на юго-западе Мексики ~9000 лет назад от своего дикого предка теосинте ( Zea mays ssp. parviglumis ) [1]. Из своего центра происхождения кукуруза испытала быстрое расширение ареала и распространилась на 90 ° широты в Америке [2, 3, 4], что потребовало новой адаптации времени цветения. ZEA CENTRORADIALIS 8 ( ZCN8 ) представляет собой ген флоригена кукурузы и играет центральную роль в опосредовании цветения [5, 6]. Здесь мы показываем, что ZCN8 лежит в основе локуса основного количественного признака (QTL) ( qDTA8 ) для времени цветения, который постоянно обнаруживается в нескольких экспериментальных популяциях кукурузы-теосинте.Путем анализа ассоциации на большой разнообразной панели инбредных линий кукурузы мы определили SNP (SNP-1245) в промоторе ZCN8 , который показал самую сильную связь со временем цветения. SNP-1245 совместно сегрегировался с qDTA8 в картографических популяциях кукурузы-теосинта. Мы демонстрируем, что SNP-1245 связан с дифференциальным связыванием активатором цветения ZmMADS1 . SNP-1245 был целью отбора во время раннего одомашнивания, что привело к почти фиксации ранее существовавшего аллеля раннего цветения в кукурузе.Интересно, что мы обнаружили независимый ассоциативный блок выше SNP-1245, в котором аллель раннего цветения, который, скорее всего, произошел от Zea mays ssp. mexicana интрогрессировал в раннецветущий гаплотип SNP-1245 и способствовал адаптации кукурузы к северным высоким широтам. Наше исследование демонстрирует, как независимые цис -регуляторные варианты гена могут быть выбраны в разное время эволюции для локальной адаптации, подчеркивая, как развиваются сложные механизмы цис -регуляторного контроля.Наконец, мы предлагаем полигенную карту доколумбового распространения кукурузы по всей Америке.
Ключевые слова
Ключевые слова
Maize
Teosinte
TEOSINTE
TEOSINTE
QTLQTL
адаптация
Выбор
DomeseStication
Рекомендуемые статьи Статьи (0)
© 2018 Elsevier Ltd.
Рекомендуемые статьи
Сообщение
Морфология цветов Zea mays на JSTOR
Перейти к основному содержанию Есть доступ к библиотеке? Войдите через свою библиотекуВесь контент Картинки
Поиск JSTOR Регистрация Вход- Поиск
- Расширенный поиск
- Изображения
- Просматривать
- По тематике
Журналы и книги - По названию
Журналы и книги - Издатели
- Коллекции
- Изображения
- По тематике
- Инструменты
- Рабочее пространство
- Анализатор текста
- Серия JSTOR Understanding
- Данные для исследований
| Штаммы и | › cv.2K1 фиолетовый › cv. 38-11 › рез. A188 › рез. A441 › рез. A554 Подробнее » › cv. A6 |